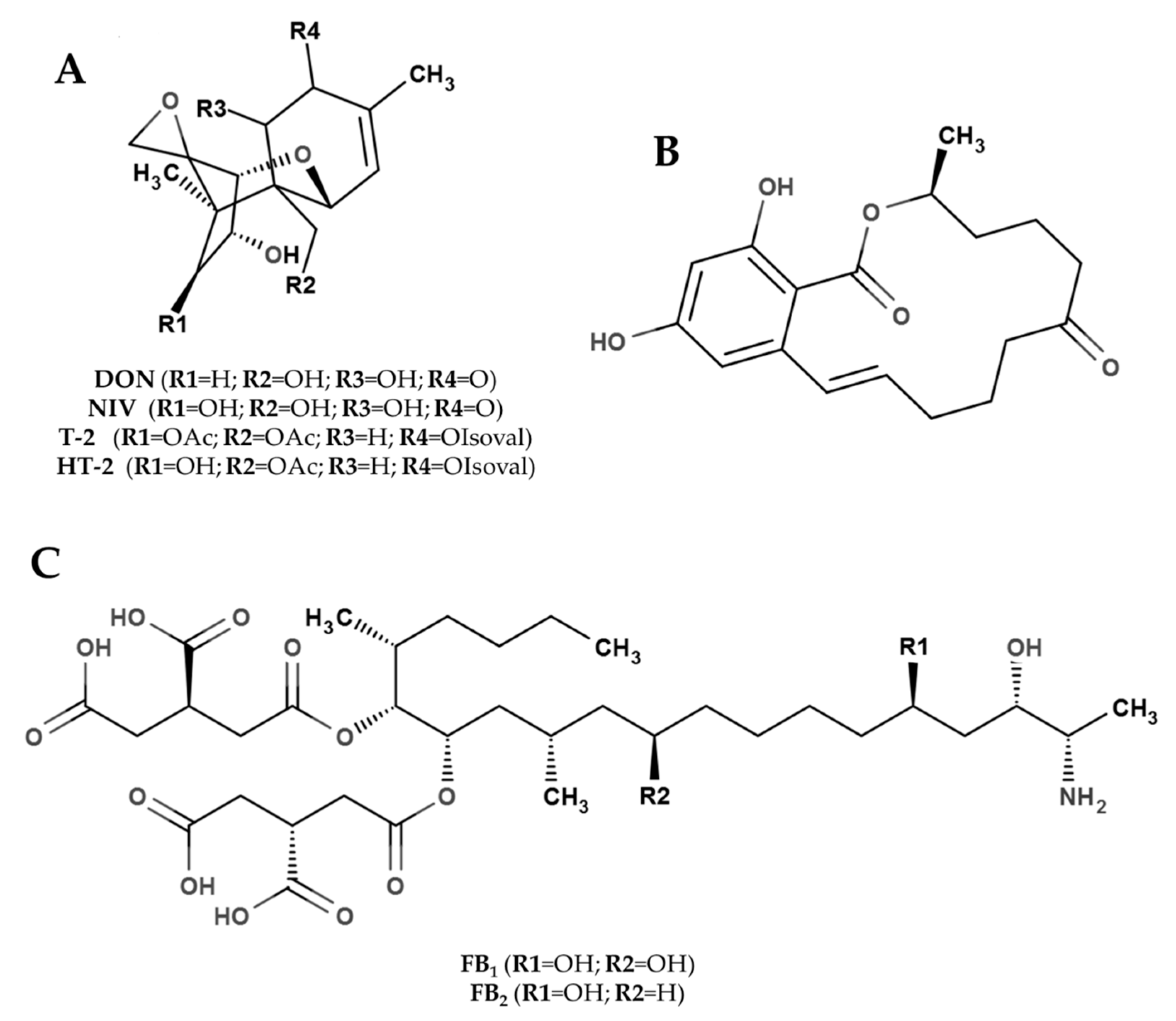
4.1. Effect of Climate Events on FHB, Maize ear Rots and Mycotoxin
Climate is among the most important factors influencing the occurrence and distribution of
Fusarium. Different climatic conditions (e.g., temperature and rainfall) in different geographical locations affect the incidence of pathogens responsible for FHB of small grain cereals and ear rots of maize. The relationships between climatic factors and FHB development have been thoroughly investigated [
14,
157]. Well define ranges of temperature and water availability are determining factors for the growth of
Fusarium and mycotoxin production [
42]. Warm and moist conditions, especially during the period of anthesis, are considered critical factors for FHB development. Among the FHB causal agents, the fungal species vary on a regional and continental scale and during any given season [
158,
159]. It is conceivable to suggest that under the influence of climatic changes, modifications in the total and relative abundance of fungal species of the FHB complex may occur. Changes in climatic extremes would have direct impacts on
Fusarium ear disease and mycotoxin production because weather factors can strongly affect epidemics and the proportions of the species responsible for FHB and ear rots [
160]. These changes could also influence the production of DON by the two main DON-producing fungal species,
F. graminearum and
F. culmorum, as well as the production of fumonisins by the main producer,
i.e.,
F. verticillioides. In fact, temperature that may be optimal for growth, are different from those optimal for mycotoxin synthesis by
F. graminearum,
F. culmorum and
F. verticillioides [
161,
162].
In maize, climatic factors determine the balances that occur within
Fusarium populations. Maize ear rots are caused by a mixture of pathogens that compete among themselves. It is generally recognized that negative interactions in competition between
Fusarium spp. are prevalent. GER and FER are favoured by distinct climatic conditions: GER is favoured by high levels of moisture at silking, followed by moderate temperatures and high rainfall during the maturation period [
163] while FER is more common in warmer and drier areas [
37]. In fact,
F. verticillioides prefers a higher temperature of 30 °C and tolerates water stress better than
F. graminearum. Fumonisin contamination is highly dependent on the composition of the
Fusarium community as well as environmental conditions, and fumonisin incidence can be high or low in relation to the growing areas. Several evidence indicates that water stress during drought events is strongly associated with high levels of
F. verticillioides infection and fumonisin accumulation in kernels [
27]. In particular, the factors that affect fumonisin development include environmental factors (temperature, humidity), insect damage and pre-/post-harvest management.
More,
F. verticillioides isolates were found to exhibit better performance at higher temperatures and under water stress conditions in comparison to
F. proliferatum, another fumonisin producing species [
164]. The levels of FB
1, the most abundant and toxicologically active fumonisin, were found to be absent or significantly low in areas generally characterized by cold and wet seasons [
165]; however, under favourable conditions in these areas, fumonisins reached significant levels [
166].
4.2. Fungal Interactions in Cereals: Consequences for Fusarium Development and Mycotoxin
Interactions among fungal species depend on biotic and abiotic factors and can play an important role in the structural organization of fungal communities. These interactions range from antagonistic to mutualistic and can be positive, negative or neutral [
167]. Through different mechanisms (competition for space and feed resources), some pathogenic species may have an advantage over other fungal species that occupy the same niche, hindering the development of less competitive fungi [
168]; on the contrary, one fungal species can improve the adaptability of other species [
169]. Therefore, the role of ecological interactions is of particular importance because these interactions can significantly affect fungal development and secondary metabolism. It is critical to take this into account to accurately assess the risk of mycotoxin contamination. Moreover, host-specific influences on intraspecific competition may dictate fungal compositions and probably mycotoxin occurrence as observed in
A. flavus populations [
170]. Several studies on interspecific interactions between only
Fusarium spp. or between
Fusarium spp. and other genera have been carried out under
in vitro conditions [
171,
172], whereas only a few have been conducted under natural conditions [
173] Negative interactions in fungal communities that occupy the same ecological niche are predominant and are based principally on competition [
174,
175].
In small grain cereals, FHB is generally associated with various fungal species, including both toxigenic (several species of
Fusarium) and non-toxigenic fungi (
Microdochium spp.), and their prevalence and abundance in the same field [
176,
177] are strongly dependent on environmental variables. Due to environmental variables under field conditions, FHB development and mycotoxin production are predicted to be more complex when more than one toxigenic species is present. Also the role of conidia has been evaluated and studies on interactions between several FHB species have shown that among
Fusaria the species producing macroconidia are the most competitive during germination [
178]. Experimental evidence supports any synergetic interactions between single isolates of
F. graminearum,
F. poae,
F. culmorum and
F. avenaceum after inoculation on wheat spikes, while in most cases, the presence of competitive interactions is more evident. In the presence of a mixed FHB infection, a large reduction in fungal biomass has been observed in comparison to single inoculations. On the contrary, mycotoxin productivity per unit of fungal biomass was found to increase dramatically in the co-inoculations, indicating that the production of trichothecene mycotoxins can be affected by competition [
173]. Some experimental studies performed in wheat do not confirm these results where intraspecies interaction appears to reduce trichothecene yield [
179]. Recent findings suggest that the behaviour of different isolates in presence of a competitor is variable mostly depending by
Fusarium strain rather than species, with a predominance of aggressive isolates [
180]. However, this study also demonstrates a lack of correlation between co-occurrence of several FHB species and an increase of
Fusarium toxins risk in wheat production.
In maize, in competition with other genera that commonly co-occur on kernels,
F. verticillioides has been found to take advantage of
Aspergillus flavus and
Penicillium spp. in mixed infections [
181,
182]. Towards other
Fusarium species, inoculation of maize with isolates of
F. verticillioides, F. proliferatum (fumonisin producers) and
F. graminearum (DON and zearalenone producer), performed under different water and temperature conditions, showed that
Fusarium populations generally decreased in presence of competitors in dependence of environmental variables. In addition, fumonisin production was generally reduced in competing interactions, whereas zearalenone was not affected and DON was increased [
183]. These
in vitro experiments indicated opposite results compared with those obtained from trials under natural conditions where
F. verticillioides has been observed to inhibit the growth of
F. graminearum [
167]. It is known that
F. verticillioides has a competitive advantage over
F. graminearum when simultaneously inoculated due to better growth and a higher spore germination rate over a wider range of temperatures and water activities [
167]. However, the impact of these interactions on mycotoxin contamination requires further investigation with respect to environmental and stress conditions. Indeed, it was demonstrated that high levels of
F. verticillioides do not necessarily result in high levels of fumonisin contamination [
184]. Insensitivity of ZEA and DON producers to competition also occurred when
F. graminearum was cultivated with
Aspergillus parasiticus, and the toxin levels were not modified [
185].
4.3. Stress Factors
Depending on their environmental growth conditions, fungi sense a variety of external signals and respond by regulating secondary metabolism [
186]. Field crops are continuously challenged by several environmental stresses that occur naturally in a certain area. Cereal growth, productivity and resistance to pathogens are closely related to environmental and agronomical input, which are both related to the response of crop plants to stress. Stress conditions imposed on develo** crops, especially during the reproductive stage, can facilitate fungal infection, mycotoxin production and grain contamination [
187].
Biotic factors such as insects, pathogens and weeds [
188,
189,
190] and abiotic factors such as hot temperatures, drought and hailstorms [
191] can affect crop physiology and productivity [
192] and may result in conditions that are favourable for mycotoxin accumulation. There is evidence that the abiotic and biotic factors that predispose plants to diseases can activate several plant responses to stress, which can indirectly influence mycotoxin production [
193]. In response to biotic and abiotic stress, plants react with a rapid and transient release of reactive oxygen species (ROS), activating a broad range of strategies to protect themselves [
194,
195,
196]. Because oxidative stress in fungi was demonstrated to modulate
in vitro the biosynthetic pathways of
Fusarium mycotoxins such as trichothecenes and fumonisins [
197,
198], it is conceivable to suggest that an alteration of the cellular redox state
in planta can affect mycotoxin accumulation.
One of the primary biotic stress factors that influence fungal colonization and mycotoxin contamination are the insects. As a consequence of phytophagous insect attack , the harmful action of insects occurs in two ways: by producing wounds that are favourable entry sites for conidia already present on the ear tissues and by causing stress conditions in plant tissues [
199] through the generation of ROS [
200]. ROS generation in plant was suggested to be a common response that persists on as long as the insect attack carries on. Although the effect of insect activity on small cereal is low, insects can still be considered a potential risk for the occurrence of FHB. In fact, pre-exposal of wheat ears to aphids as
Rhopalosiphum padi and
Sitobion avenae, can co-occur with FHB appearance and lead to a significant increase in
F. graminearum colonization and DON accumulation [
201,
202]. These results are probably related to the elicitation of defense signalling pathways through accumulation of H
2O
2 and ROS [
203] as well as enhancement of plant defence [
202]. As regards to the redox potential, this can act as a modulator of DON biosynthesis [
204] that, in turn, lead to a further accumulation of H
2O
2 in wheat tissues [
205]. Recently, in
F. graminearum the gene FGK3, recognized as an important virulence factor essential for pathogenicity and DON production, was demonstrated to be up-regulated in response to H
2O
2, cold and SDS stresses [
206]. Concerning plant defence, the pathogen can produce more DON in an attempt to circumvent the enhanced defences, with the consequent acceleration of disease progression and mycotoxin accumulation [
207].
In maize,
F. verticillioides infection is facilitated by insect damage while
F. graminearum mainly infect ear through the silks, therefore, the effect of insect stress is higher for FER pathogens respect to GER ones [
5]. Recent evidence has also shown a correlation between ear-feeding insects and mycotoxin contamination in maize [
193,
208,
209], and other findings suggest that kernel-feeding insects are more important than silk- or cob-feeding insects [
210]. Insects affecting maize such as
Ostrinia nubilalis (European corn borer; ECB),
Sesamia nonagrioides (Mediterranean corn borer),
Helicoverpa zea (Corn earworm), and
Sitotroga cerealella (Angoumois grain moth) can produce tunnels into stalks and ears and can carry
F. verticillioides conidia and therefore transmit infection. Beyond to corn borers, also populations of ear-feeding insects as
Frankliniella occidentalis (Western flower thrips) provide inoculum sites for
Fusarium spp. and their presence are strongly correlated with disease severity and fumonisin contamination [
24].
While it is difficult to distinguish the role of these actions in the fungal infection process and mycotoxin induction, at least for aflatoxin accumulation, it was reported that
Aspergillus flavus infection mediated by a vector was more conducive than that mechanically mediated, demonstrating the importance of insects in mycotoxin occurrence [
211]. Because of the increased level of ROS reported after insect attack on Lima bean and potato [
212,
213] also the interaction between maize and ECB should represent a stress condition; however, how this stress relates to mycotoxin induction requires further investigation.
The results from several studies suggest a role of ROS such as H
2O
2 in mycotoxin production by toxigenic
Fusarium, as well as antioxidant compounds have been demonstrated to inhibit toxinogenesis [
198,
214]. The concomitance of multiple pathogens can positively influence
Fusarium disease in cereals. In maize, the infection by ear-damaging pathogens as
F. graminearum and
F. subglutinans facilitates the subsequent
F. verticillioides infection and fumonisin accumulation [
215]. Further evidences are provided by infection of maize with
Ustilago maydis where fumonisin levels resulted increased in the kernels harvested from smutted ears compared with the kernels from smut-free control ears [
189]. Together, these data show that an initial infection can breach the host defense and weaken plants, allowing access to other pathogens, including toxigenic fungi, and promoting their performance.
Weeds represent a threat to the crop and also an indirect stress affecting the crop performance. As reported in maize, light competition with the perennial ryegrass
Lolium perenne unfavourably modifies the pattern of plant growth and development. This interaction also highlighted as a first stress due to shade avoidance may affect sensitivity toward a subsequent abiotic stress [
216]. Moreover, the light reflected from the tissues of the above-ground neighbouring weeds was found to reduce total root biomass [
190], furtherly influencing ability of plant to adsorb water and nutrients. Finally, competition between crop plants and weeds for water, nutrients and sunlight involves the ability of a plant to respond to diseases and parasites and crop competition with weeds is presumed to increase sensitivity to soil-borne mutualists and pathogens [
217].
Abiotic stress, such as hot temperatures and drought conditions, strongly alters the efficiency of photosystems and the stability of membranes, and is associated with oxidative stress in plants [
196]. In fact, crop resistance to stress conditions can be related to high efficiency ROS-scavenging systems, as has been reported in wheat [
218]. Because dry conditions typically accompany excessive heat, it is difficult to determine the influence of single factors. During kernel filling, drought and high temperature are considered as the environmental conditions that are most conducive to mycotoxin contamination in maize [
219]. In Poland, it was reported that particularly high concentrations of fumonisins were associated with the hottest and driest summers [
220]; however, other studies reported a low [
24] or no influence of drought stress [
221]. Moreover, during field trials conducted in Italy under climatic conditions that were considered unfavourable for
Fusarium infections, a decrease in
F. verticillioides colonization in maize was not related to an equal reduction in fumonisin accumulation. This result further suggests that hot conditions and drought stress play an important role in modulation of fumonisin production [
184]. Environmentally damaging conditions such as hailstorms have also been reported to decrease quality [
222] and increase mycotoxin contamination [
191], favouring the entry of a fungal pathogen and causing plant stress. These authors reported that fumonisins were more frequently detected in grain from hail-damaged fields compared with undamaged fields. While an increase in the level of stress signalling following mechanical damage of leaf tissues has been well documented [
223], the possibility that this type of meteorological event can stimulate mycotoxin biosynthesis through wound signals should be considered.





{kind=link}