1. Introduction
Cucumber is one of the most important vegetables grown in the world, especially in China. Fusarium wilt, a soil-borne disease caused by
F. oxysporum, and anthracnose, which is caused by
C. lagenarium, are the main diseases that limit the yield and quality of cucumber worldwide [
1,
2]. Stems and roots, and especially root wounds, can be infected by
F. oxysporum. The chlamydospores generated by
F. oxysporum can survive in infested field soil for several years [
3]. On the other hand, all above-ground tissues of cucumber, such as leaves, stems, petioles, and fruits, can be infected by
C. lagenarium [
4].
In practice, the application of chemical fungicides is still the main method used for disease management in cucumber due to the limit of highly resistant cultivars [
5,
6]. However, widespread use of these chemical fungicides has generated serious problems, including cost, environmental pollution, decreased efficacy due to pathogen resistance, and threats to human health [
7,
8]. Therefore, screening for alternative fungicides that do not harm people or the environment is urgently necessary. Considering the requirements of modern greenhouse agriculture, botanical fungicides have attracted more attention. Indeed, the practice of using botanical fungicides, or plant extracts as we know them in agriculture, dates back at least two millennia in many countries, such as ancient China, Greece, India, Egypt, and North America [
9]. Botanical fungicides have certain advantages: They usually break down rapidly and do not persist in the environment, and have a low risk of inducing pathogen resistance. Moreover, botanical fungicides can also be used as lead compounds for the synthesis of novel compounds [
10,
11].
Cuminic acid was extracted from the seed of
Cuminum cyminum L. in our previous research [
12]. Cuminic acid belongs to the benzoic acid chemical group. Previous studies have shown that cuminic acid exhibits obvious antifungal activity against several plant pathogens, such as
Phytophthora capsici Leonian,
Rhizoctonia cerealis E.P. van der Hoeven, and
Sclerotinia sclerotiorum (Lib.) de Bary. The mycelial growth of
S. sclerotiorum and
R. cerealis was completely inhibited when treated with cuminic acid at 200 μg/mL [
13,
14]. Moreover, in greenhouse experiments, more than 50% efficacy against
S. sclerotiorum and
Blumeria graminis f. sp.
tritici was obtained when plants were treated with cuminic acid at 1000 μg/mL, which was comparable with the efficacy of procymidone at 100 μg/mL [
13]. Moreover, the EC
50 value of cuminic acid for inhibiting the mycelial growth of
P. capsici was only 14.54 µg/mL, which was even lower than the EC
50 value of the natural compound eugenol against
P. capsici [
12].
Cuminic acid has demonstrated broad-spectrum antifungal activity. However, few reports about the antifungal activity of cuminic acid against F. oxysporum or C. lagenarium are available in the literature. Therefore, the aim of this study was to evaluate the potential value of the natural compound cuminic acid, extracted from the seed of Cuminum cyminum L., as a botanical fungicide against F. oxysporum or C. lagenarium.
3. Discussion
Plant diseases Fusarium wilt and anthracnose, caused by pathogenic fungi, are the most important factors that limit cucumber production worldwide, especially in China. In practice, most efforts to control the two diseases have focused on the application of synthetic chemical fungicides [
5,
6]. However, serious problems, such as handling hazards, fungicide resistance, and threats to human health and the environment, have generated concern. Therefore, looking to botanical fungicides or plant extracts has started in an attempt to find alternatives.
In our previous study, 1.2 g cuminic acid was extracted from 15 kg of
Cuminum cyminum L. seed [
15]. Importantly, the toxicity test of 1% cuminic acid micro-emulsion on animals and plants, such as quail, bee, silkworm, carp, earthworm, tadpole, wheat, chili, and oilseed rape, was performed. The results were as follows: the LD
50 for quail was greater than 1000 mg/kg; for bees it was greater than 5000 mg/L; for silkworm it was greater than 1500 mg/L; for carp it was greater than 1600 mg/L; for earthworm it was greater than 1000 mg/L; for tadpoles it was greater than 40 mg/L; and it exhibited low risk for wheat, chili, and oilseed rape [
16]. All the results suggested that cuminic acid was safe and would not biologically target any sensitive species.
In the current study, the antifungal activity of cuminic acid, which was extracted from the seed of
Cuminum cyminum L., was assessed. The results showed that cuminic acid has obvious inhibitory activity against
F. oxysporum and
C. lagenarium. Moreover, cuminic acid was more active against spore germination than mycelial growth, which was consistent with its activity against
P. capsici [
12]. However, there was no significant difference among the EC
50 values for carbendazim against mycelial growth or spore germination, indicating that the mechanism of action was different between cuminic acid and carbendazim.
In a pathogen’s lifecycle, spore germination and mycelial growth are two different stages. Once infection has occurred, the suppression of mycelial growth within the host tissue becomes a major disease management target, which could interrupt the lifecycle of the pathogen [
17]. In greenhouse experiments, cuminic acid at 2000 µg/mL exhibited over 60% protective and curative activity against
F. oxysporum, which was nearly equal to the efficacy obtained by the chemical fungicide carbendazim, indicating that cuminic acid has both protective and curative activity. Therefore, it not only reduced the infection ability of the mycelia but also inhibited the spore germination. Cuminic acid was also detected in both the roots and leaves using HPLC, indicating that cuminic acid exhibits good characteristics of absorption and can be transported both upward and downward in cucumber. Cuminic acid could be used for the control of both foliar disease and root disease, which is different compared to the previous study that showed that carabrone could be only transported upward instead of downward [
18].
Previous studies have demonstrated that plants have special immune sensors that can identify bacteria, viruses, fungi, and other microbial invasion. The enzyme system in plants can not only regulate the normal physiological activities, but also prevent the invasion of microorganisms [
19,
20]. In addition, salicylic acid and jasmonic acid are two important signal transmission compounds that induce plants’ defense systems [
21,
22]. In this study, the POD and CAT activity of the cucumber leaves decreased after being treated with cuminic acid, whereas the SOD and PPO activity increased significantly after being treated with cuminic acid. Work to explore the expression of genes correlated with the pathway of salicylic acid or jasmonic acid, which is associated with the plant’s defense ability, is underway in our lab.
Integrated control of fungal diseases, including biological controls and safer chemicals such as food preservatives, chitosan, and plant-derived products, has been studied [
23,
24]. Benzoic acid has been commonly used as food preservative and is widespread in industrial wastewater [
25]. Cuminic acid, extracted from the seed of
Cuminum cyminum L., belongs to the benzoic acid chemical group. However, a previous study demonstrated that cuminic acid did not target organisms, was easily degradable in soil and water, and was environmentally friendly [
26].
Over several decades, crop growers have generally apply synthetic fungicides as the main method for controlling plant diseases. However, modern organic agriculture, where products cultivated without applying any chemical pesticides, is increasing in popularity [
27]. Natural plant chemicals, which are the major sources of industrial and medicinal materials, including plant extracts and essential oils, have also shown potential for agricultural pest management [
28,
29]. For example,
Vitex agnus-castus extract exhibited strong antifungal activity against
Pythium ultimum in tomatoes under both in vitro and in vivo conditions [
30];
Cortex Pseudolaricis extract exhibited potential antifungal activity against
Colletotrichum gloeosporioides [
31]; and the essential oil from
Tetradium glabrifolium fruits showed larvicidal activity against
Aedes albopictus [
32]. In addition, glucosinolates, terpenoids, flavor compounds, and glucosinolates are receiving increased attention worldwide [
33]. Previous studies demonstrated that natural products could not only be directly used as crop protection agents, but could also be used as lead compounds for the synthesis of new pesticides, such as a series of strobilurin fungicides, including azoxystrobin and picoxystrobin, which were derived based on the structure of strobilurin A [
34,
35,
36,
37]. Considering the public concern about the impact of synthetic pesticides on human health and the environment, natural products extracted from microbes, plants, and other organisms will continue to be important sources for environmentally friendlier pest management tools.
4. Materials and Methods
4.1. Fungicides and Strains
Cuminic acid (98%) was purchased from Aladdin Bio-Chem Technology Company (Shanghai, China) and dissolved in 10 mL methanol to 100 mg/mL for stock solutions. Carbendazim (98%), provided by Shenyang Study Institute of Chemical Industry (Shenyang, China), was dissolved in 0.1 mol/L hydrochloric acid (HCl) at 10 mg/mL as stock solutions.
Five F. oxysporum strains and four C. lagenarium strains (single-spore isolates) were provided by the Research and Development Center of Biorational Pesticides, Northwest A & F University and maintained on potato dextrose agar (PDA) slants at 4 °C.
4.2. Sensitivity to Cuminic Acid
The EC
50 values of cuminic acid for the inhibition of mycelial growth were determined according to a previous study [
12]. Inverted mycelia plugs, 5 mm in diameter, were cut from the margin of the 5-day-old colonies and then transferred to PDA plates which were amended with cuminic acid at concentrations of 0, 6.25, 12.5, 25, 50 and 100 μg/mL. After the plates were incubated at 25 °C for 5 days, colony diameters in two perpendicular directions were measured and averaged. The inhibition rate of mycelial growth was calculated [
12].
The activity of cuminic acid on the inhibition of spore germination was determined as per a prior study [
37]. The strains were first incubated on PDA plates in the dark at 25 °C for 7 days. Then, the plates were drenched with 15 mL of sterile distilled water, and the spores were carefully collected from the culture surface. Spore suspensions for each strain, prepared at 1 × 10
5, were spread on water agar plates amended with cuminic acid at 0, 3.125, 6.25, 12.5, 25 and 50 μg/mL. After incubation at 25 °C for 10 h, the germination of conidia was checked and the inhibition rate of spore germination was calculated. In terms of sensitivity, carbendazim was used as the control fungicide and the concentrations of carbendazim used were 0, 0.3125, 0.625, 1.25, 2.5, 5, 10, and 20 μg/mL. The experiment was conducted three times with three replicates per treatment.
4.3. Protective and Curative Activity of Cuminic Acid in Greenhouse Experiments
The protective activity of cuminic acid against
F. oxysporum or
C. lagenarium was conducted as follows [
30]. One cucumber plant per pot, at a similar growth stage (three to five leaves), were irrigated with 10 mL of water, and carbendazim at 500 μg/mL or cuminic acid at 500, 1000 or 2000 μg/mL. After 24 h, cucumber plants were irrigated with 10 mL of spore suspension (1 × 10
5 spores) collected from
F. oxysporum, or sprayed with 10 mL of spore suspension (1 × 10
5 spores) collected from
C. lagenarium.
Then, the irrigated plants were kept at 25 °C with 80% humidity in a growth chamber. All leaves from each plant and six plants for each concentration were used for the detection of anthracnose. The experiment was repeated three times. The disease indeices of Fusarium wilt and anthracnose were detected 15 days or 5 days after inoculation, respectively, and the control efficacy was calculated [
8,
38].
A 0–4 visual scale grade of the rhizomes and roots was used to determine the disease severity of
F. oxysporum, where 0 represented rhizomes and roots with no symptoms, 1 denoted lesions on less than 25% of the total area, 2 denoted lesions on 25–50%, 3 denoted lesions on 50–75%, and 4 represented lesions on more than 75%. The disease index was calculated as 100 × [(n × 1 + n × 2 + n × 3 + n × 4)]/(4× total assessed plants), where n represents the number of diseased plants in that grade [
38].
A 0–9 disease severity grade was used to test the disease severity of
C. lagenarium, which was evaluated visually on individual leaves as the percentage of diseased area. 0 represented no symptoms; 1 indicated <1%, 3 indicated 1–10%, 5 indicated 10–25%, 7 indicated 25–50%, and 9 indicated >50%. The disease index was calculated as 100 × [(n × 1 + n × 3 + n × 5 + n × 7 + n × 9)]/(9 × total number of examined leaves), where n represents the numbers of leaves corresponding to the disease grade [
8].
The equation to calculate control efficiency was:
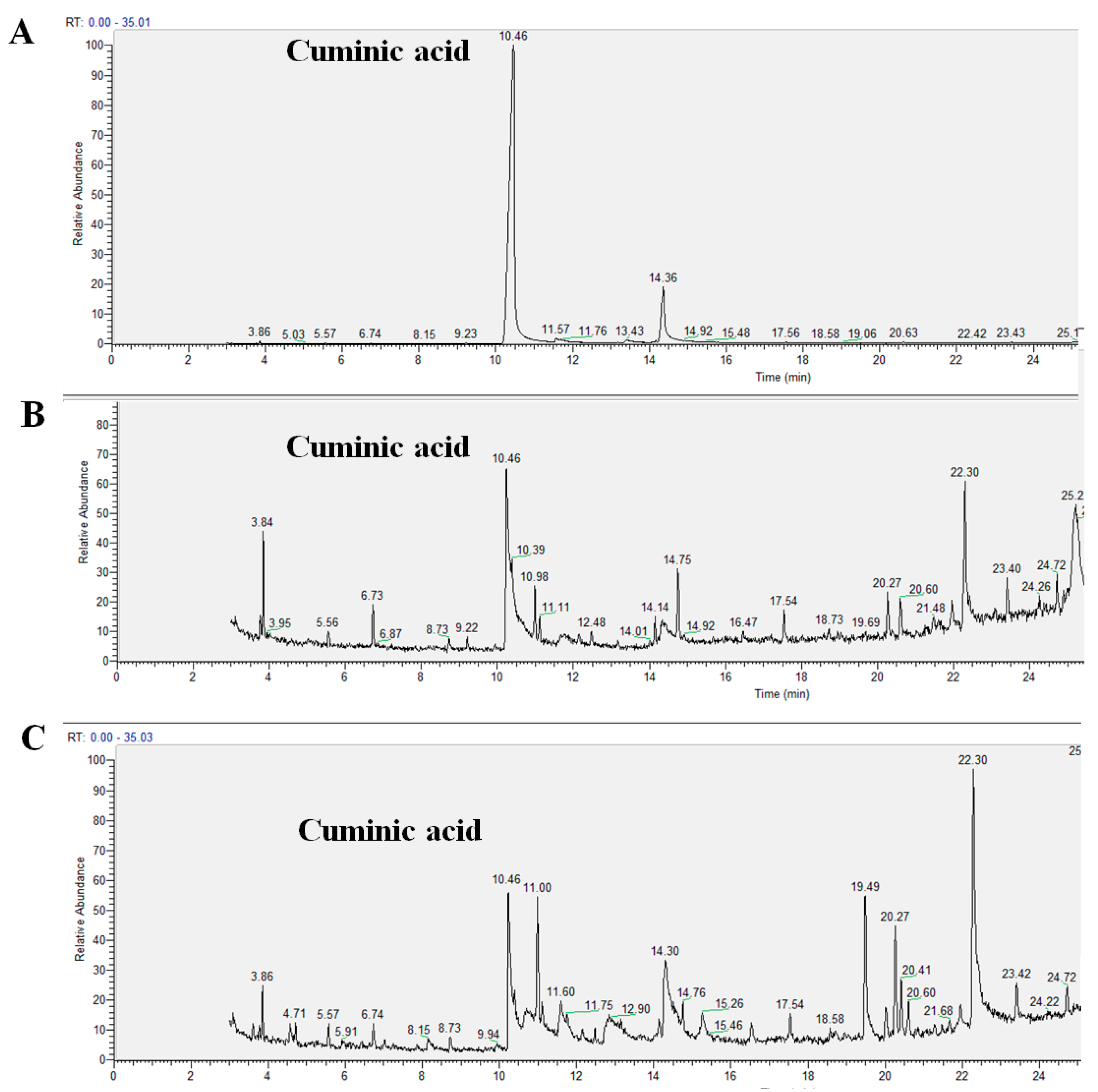
4.4. Translocation of Cuminic Acid in Cucumber Plants
To detect the downward translocation of cuminic acid, leaves of cucumber plants were sprayed with cuminic acid at 500 µg/mL until runoff. After the plants were incubated in a greenhouse for 3 days, rot of the plants were cut off and ground in liquid nitrogen. After being mixed with 5 mL of methanol, the solution was detected using high performance liquid chromatography (Waters Alliance 2690, Waters, Milford, MA, USA) [
17]. For detecting the upward translocation of cuminic acid, cucumber plants were irrigated with 10 mL cuminic acid at 500 µg/mL. Then, the leaves were cut off and treated as described above. Cuminic acid (98%) was used as the standard product. The experiment was conducted twice with four plants per treatment.
HPLC detection conditions were as follows: Hpersil BDS-C18 (4.6 mm × 250 mm, 10 mm, Thermo Fisher Scientific, Waltham, MA, USA) was used. Seventy percent methanol was used as the mobile phase and the flow rate was 1 mL/min with an injection volume of 10 µL. The detection wavelength was 233 nm and the column was kept at room temperature.
4.5. Superoxide Dismutase, Peroxidase, Polyphenol Oxidase, and Catalase Activity
All the leaves of cucumber plants at a similar growth stage were sprayed with water and cuminic acid at 500 µg/mL until runoff and then kept at 25 °C with 80% humidity. The plants were not inoculated with the pathogens. After incubation for 1, 3, 5 and 7 days, one leaf per plant was cut off (0.2 g per leaf) and the superoxide dismutase (SOD), peroxidase (POD), polyphenol oxidase (PPO), and catalase (CAT) activities were determined using commercial kits (Jiancheng, Nan**g, China) according to the manufacturer’s instructions. Six plants per treatment were used and the experiment was repeated three times.
4.6. Data Analysis
Due to the variances between experiments being homogeneous, data from repeated experiments were combined for analysis. Statistical analysis was conducted using SPSS 14.0 (SPSS Inc., Chicago, IL, USA). The EC50 values of the strains were calculated by linear regression of the log of the colony diameter versus fungicide concentration. The ANOVA procedure of SPSS and Fisher’s protected least significant difference (p = 0.05) were used to determine whether significant differences existed among the data.






{kind=link}
{kind=link}
{kind=link}
{kind=link}