1. Introduction
Plants are frequently threatened by a variety of biotic and abiotic stresses during their life cycle. Salt-alkaline soils are widely distributed across arid and semi-arid regions of the world, and have detrimental effects on plant growth and development [
1,
2]. Although soil alkalization is often associated with soil salinity, the former is considered much more hazardous to plants. This condition is generally linked with high pH stress and sodium toxicity caused by an excess of Na
2CO
3 and NaHCO
3 in the soil, as well as osmotic stress. The comprehensive stress caused by alkaline soils directly affects physiological homeostasis at the cellular and whole-plant levels [
3,
4,
5]. How plants respond to salt stress has been widely studied; for example, the SOS (Salt Overly Sensitive) pathway has been identified as one of the most important signaling pathways established in plants [
6,
7]. The vacuole-bound NHX-like Na
+/H
+ antiporters also contribute to the plant response to salt stress [
8]. In contrast, less attention has been given to the mechanism by which plants respond to alkaline stress.
Plants develop various physiological and biochemical strategies to cope with stresses. Reactive oxygen species (ROS) are produced as a byproduct of several cell processes and can act as positive modulators similar to the role of phytohormones [
9,
10,
11]. However, negative environmental stimuli can lead to excess ROS accumulation and irreversible damage, e.g., degradation of chlorophyll, lipid peroxidation, and deterioration of nucleic acids [
12,
13]. Although the plant response to stresses can involve different physiological and molecular changes, the accumulation of ROS is a common reaction. Therefore, plants mobilize sophisticated antioxidant systems (both enzymatic and non-enzymatic) to maintain tight control of ROS homeostasis [
14,
15]. For example, superoxide dismutase (SOD) is the only enzyme in plant cells that can convert excess superoxide anion (O
2−) into hydrogen peroxide (H
2O
2), while both catalase (CAT) and peroxidase III (POD) function to convert H
2O
2 into water and oxygen [
14,
16].
Scavenging of excess ROS is a direct strategy by which plants adapt to adverse environments. However, plants can also alter their metabolism and accumulate beneficial metabolites, including polyamines (PAs). These small, flexible, nitrogen-containing compounds are found in almost all living cells, with the most common PAs in higher plants being putrescine (Put), spermidine (Spd), and spermine (Spm) [
17]. Changes in polyamine (PA) metabolism are positively correlated with plant tolerance to stresses such as drought, salinity, cold, and pathogens, as well as modifications in the expression of PA biosynthetic genes [
18]. For example, FcWRKY70 positively regulated Put synthesis to confer drought tolerance in
Fortunella crassifolia [
19]. Spd promotes biomass accumulation and upregulates proteins involved in cell rescue and defense as well as antioxidant enzymes in tomato (
Lycopersicon esculentum) seedlings under high-temperature stress [
20]. Exogenous Spm was proved to induce defense response and increase resistance to root rot pathogen
Phytophthora capsici in
Capsicum annuum plants [
21]. Furthermore, a study in
Oryza sativa L. demonstrated that Spd is vital for the adjustment of intracellular PA pathways and endogenous PA homeostasis that promotes salt tolerance [
22].
Melatonin, a small, flexible, and highly conserved molecule with an indole structure, is ubiquitous in a wide range of cells, from bacterial to mammalian [
23]. Since it was first reported in plants in 1995, melatonin has been shown to regulate fundamental cellular and physiological processes, e.g., stress responses [
24,
25]. In many plant species, melatonin confers enhanced tolerance during their responses to environmental stimuli such as salt, drought, extreme temperatures, heavy metals, or UV damage [
26,
27,
28,
29,
30]. The beneficial role of melatonin in sodic alkaline stress response has also been mentioned in tomatoes [
31,
32]. However, whether melatonin contributes to alkaline stress in perennial woody plants remains unclear. This molecule is thought to have direct and indirect antioxidant activity [
33,
34,
35]. Proteomics analysis of
Cucumis sativus has indicated that melatonin facilitates ATP production as an energy source for seedlings exposed to high salinity [
36]. Plants of
Medicago sativa that are pre-treated with melatonin show better regulation of proline metabolism and nitro-oxidative homeostasis, which improves their degree of drought tolerance [
28]. Under stress conditions, melatonin also strengthens several metabolic pathways, such as polyamine metabolism, ribosome pathway, amino acid metabolism, and so on [
37].
As described here, we utilized a hydroponics system to simulate alkaline conditions and investigated whether exogenous melatonin could enhance stress tolerance in seedlings of Malus hupehensis Rehd. In particular, we focused on the potential antioxidative properties of melatonin, and examined its roles in PA metabolism and the expression of genes involved in PA synthesis.
3. Discussion
Soil alkalization is a major abiotic stress that impairs plant growth and productivity worldwide. Saline-alkaline soils cover approximately 30% of the world’s land; in northeastern and northwestern China, these conditions continue to expand, posing a serious threat to agriculture [
38,
39]. Soil alkalization is coupled with soil salinity. Although many pathways have been elucidated that regulate plant responses to salt stress, fewer studies have focused on the mechanisms by which plants, especially perennial fruit trees, adapt to alkaline conditions. Our results indicated that exogenous melatonin enhances tolerance to alkaline stress in
Malus hupehensis Rehd. by regulating the biosynthesis of polyamines.
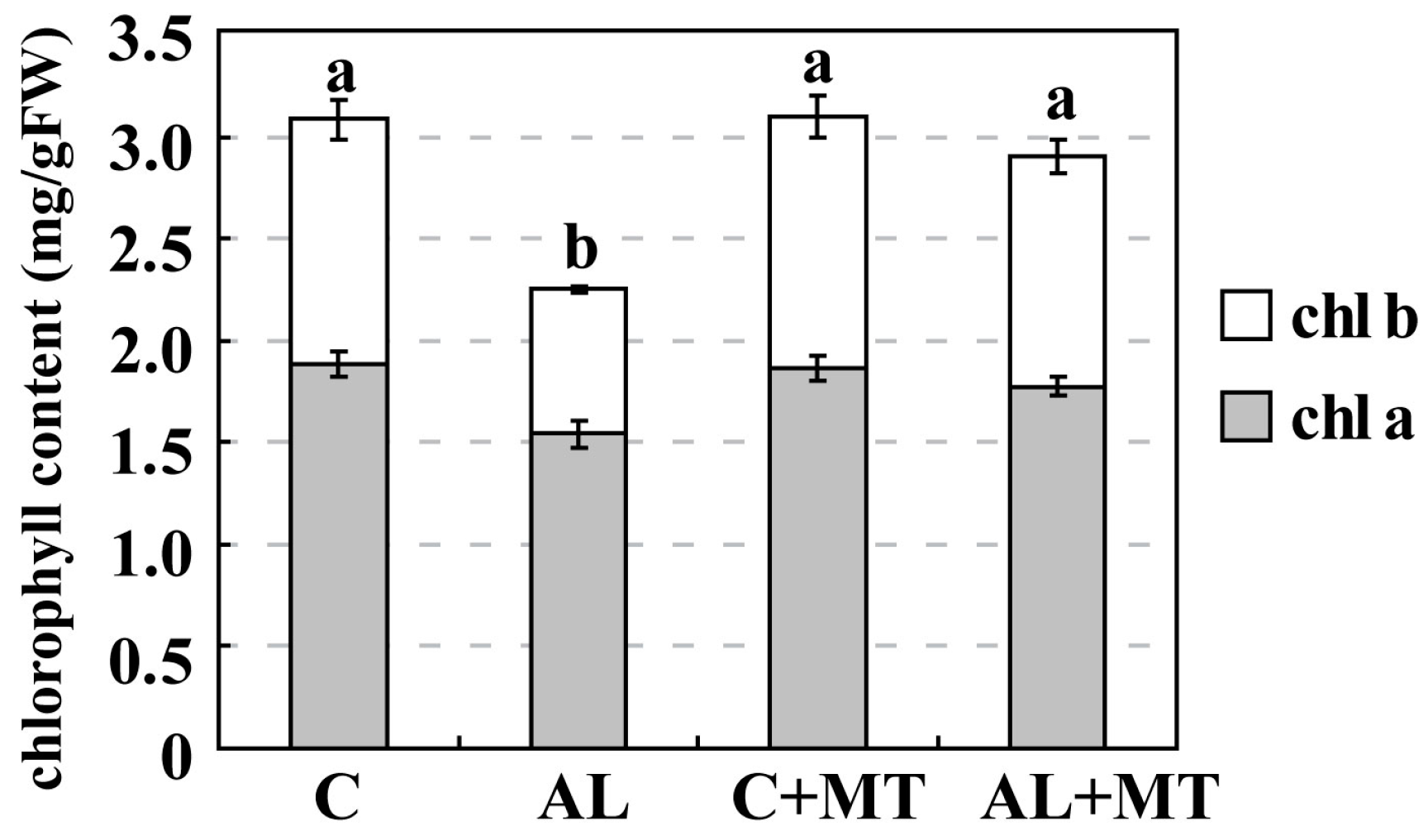
The foliage was chlorotic and growth was retarded in response to alkaline stress. After 15 days of treatment, those seedlings were also shorter than the control and had produced very little new biomass. In
Arabidopsis plants, endogenous melatonin levels are induced by heat stress and exogenous melatonin improves their thermotolerance [
40]. When the sheep serotonin
N-acetyltransferase gene
NAT is expressed in rice (
Oryza sativa cv. Dong**), those plants produce more melatonin when compared with the wild type, and the transgenics also exhibit resistance to herbicide-induced oxidative stress [
41]. Supplementing
Hordeum vulgare plants with exogenous melatonin causes them to accumulate more ABA and enhances their tolerance to cold stress [
27]. Melatonin also confers salt tolerance in watermelon (
Citrullus lanatus L.),
Malus hupehensis Rehd., and
Helianthus annuus [
26,
42,
43]. The negative effects of alkaline stress are somewhat similar to those of salt stress. We found here that, under alkaline stress,
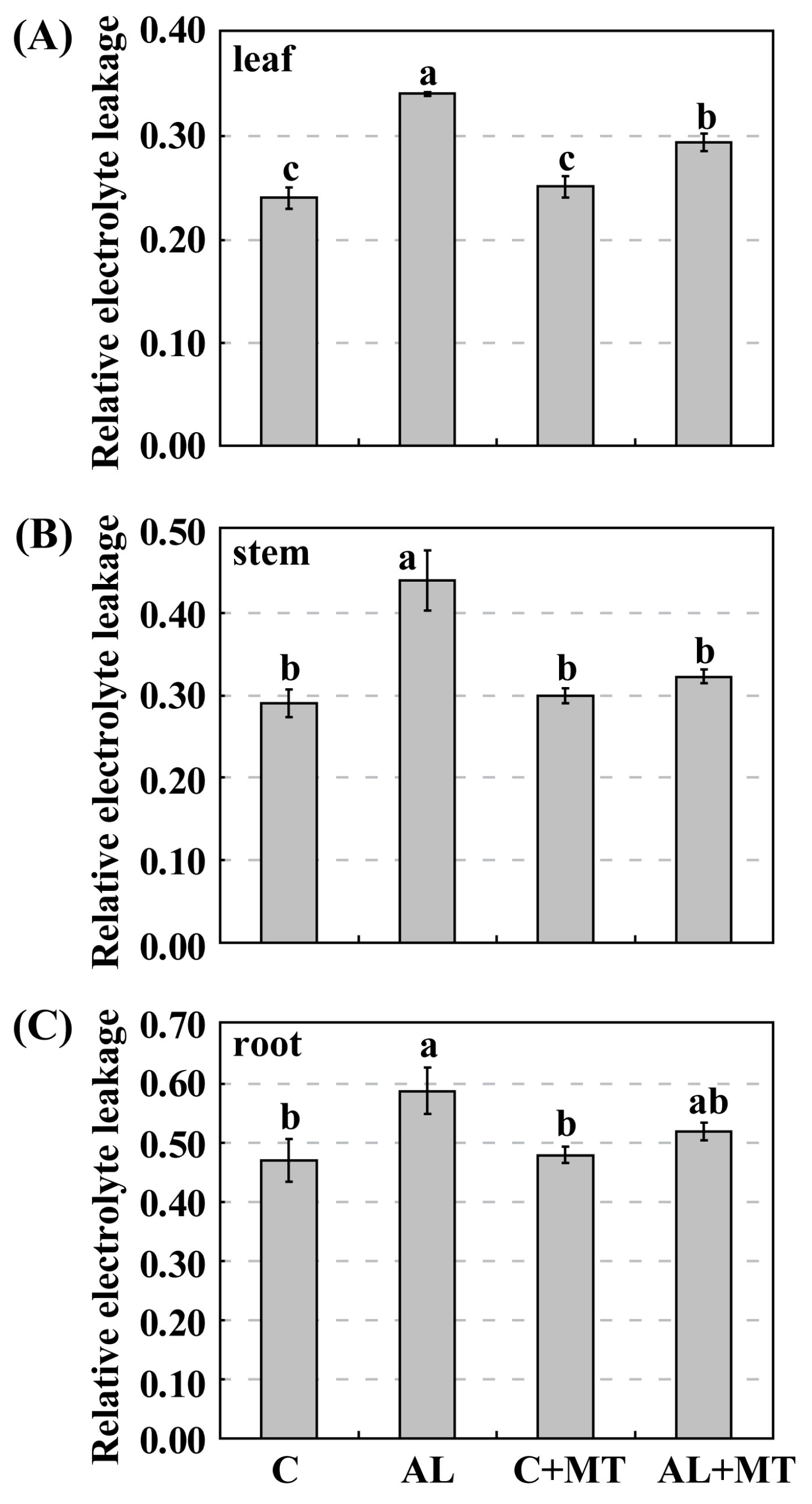
M. hupehensis seedlings that had been pre-treated with exogenous melatonin grew taller and were stronger than those that had not received that supplement. They were not as chlorotic, based on their higher chlorophyll concentrations, and showed less damage to their cell membranes, as evidenced by their relatively lower REL values. All of these findings, as well as those reported earlier, demonstrated that melatonin is a positive regulator of plant stress responses, and that exogenous applications improve plant tolerance to alkaline stress.
Plants require a healthy root system for water absorption and nutrient uptake. In responding to a changing soil environment, plants can modify various aspects of their root system architecture (RSA), including root length, branching, and total surface area [
44,
45]. When the supply of water and nitrates fluctuates, plants of
Arabidopsis show a change in the lengths of their primary roots and basal root growth [
46]. In rice, DEEPER ROOTING 1 (DRO1) is a quantitative trait locus that controls root angle. Its expression in that crop leads to deep rooting and helps plants avoid the negative effects of drought [
47]. Exposing rice seedlings to a saline-alkaline medium results in significant reductions in the number of adventitious roots, total root length, and root surface area [
48]. Under normal growing conditions, plants of
Lotus japonicus display a dichotomous RSA pattern that becomes a herringbone pattern in response to alkaline stress [
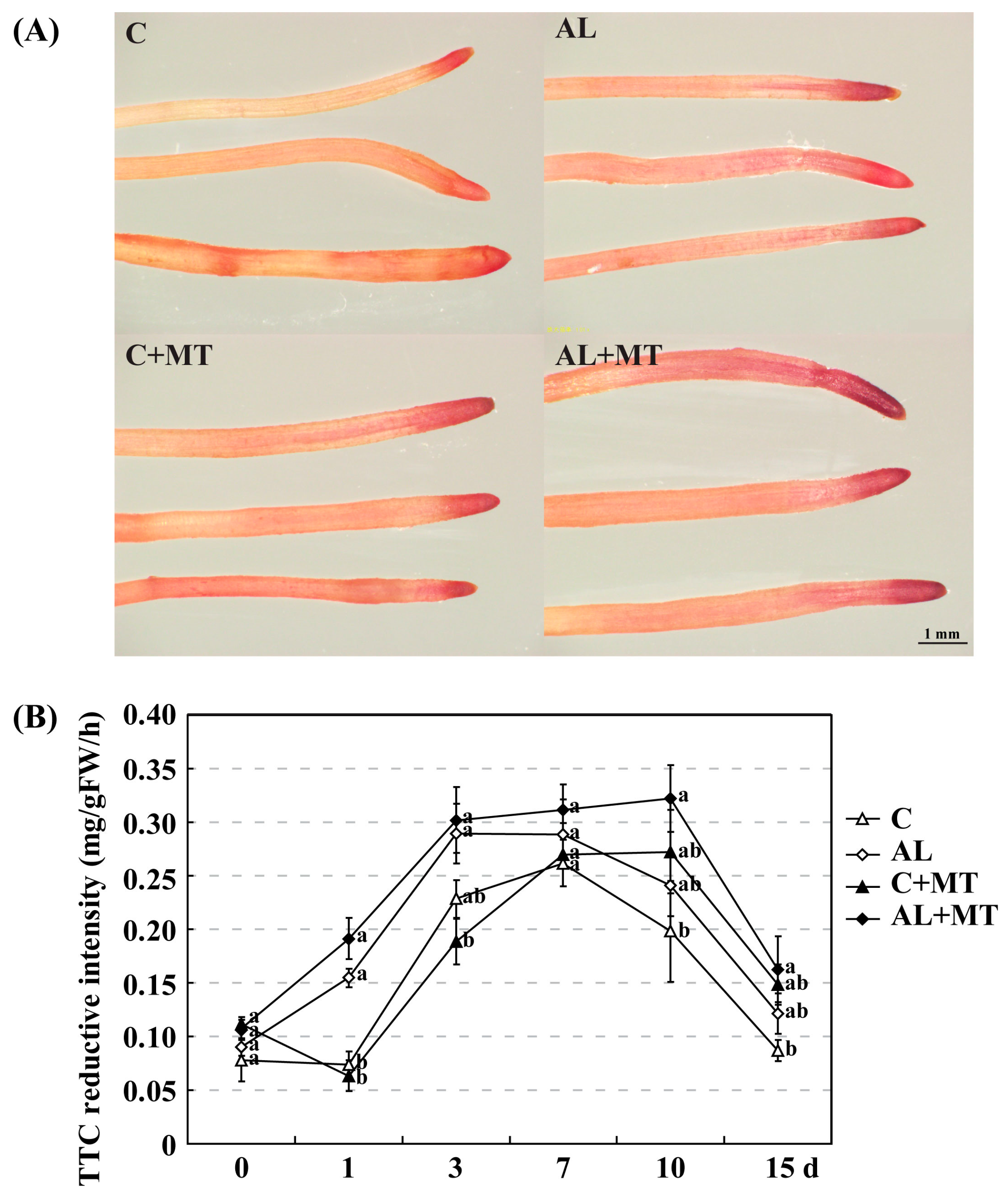
49]. For our seedlings, the values recorded for all six RSA parameters—root length, surface area, average diameter, volume, and numbers of tips and forks—were the lowest under stress conditions. In contrast, treatment with melatonin promoted the development of a strong RSA by modifying specific physiological processes in the root cells, and increased root activity in stressed seedlings.
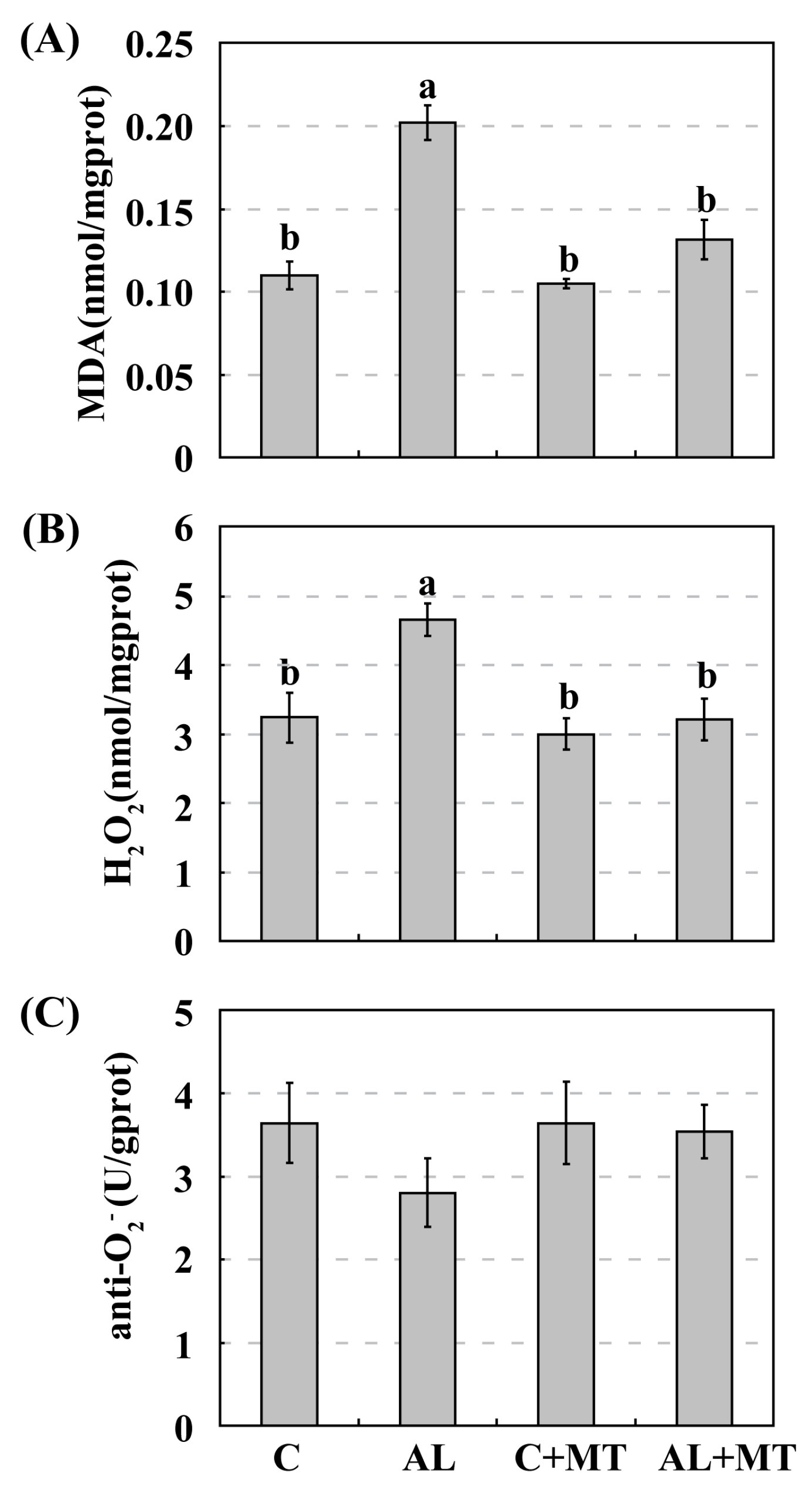
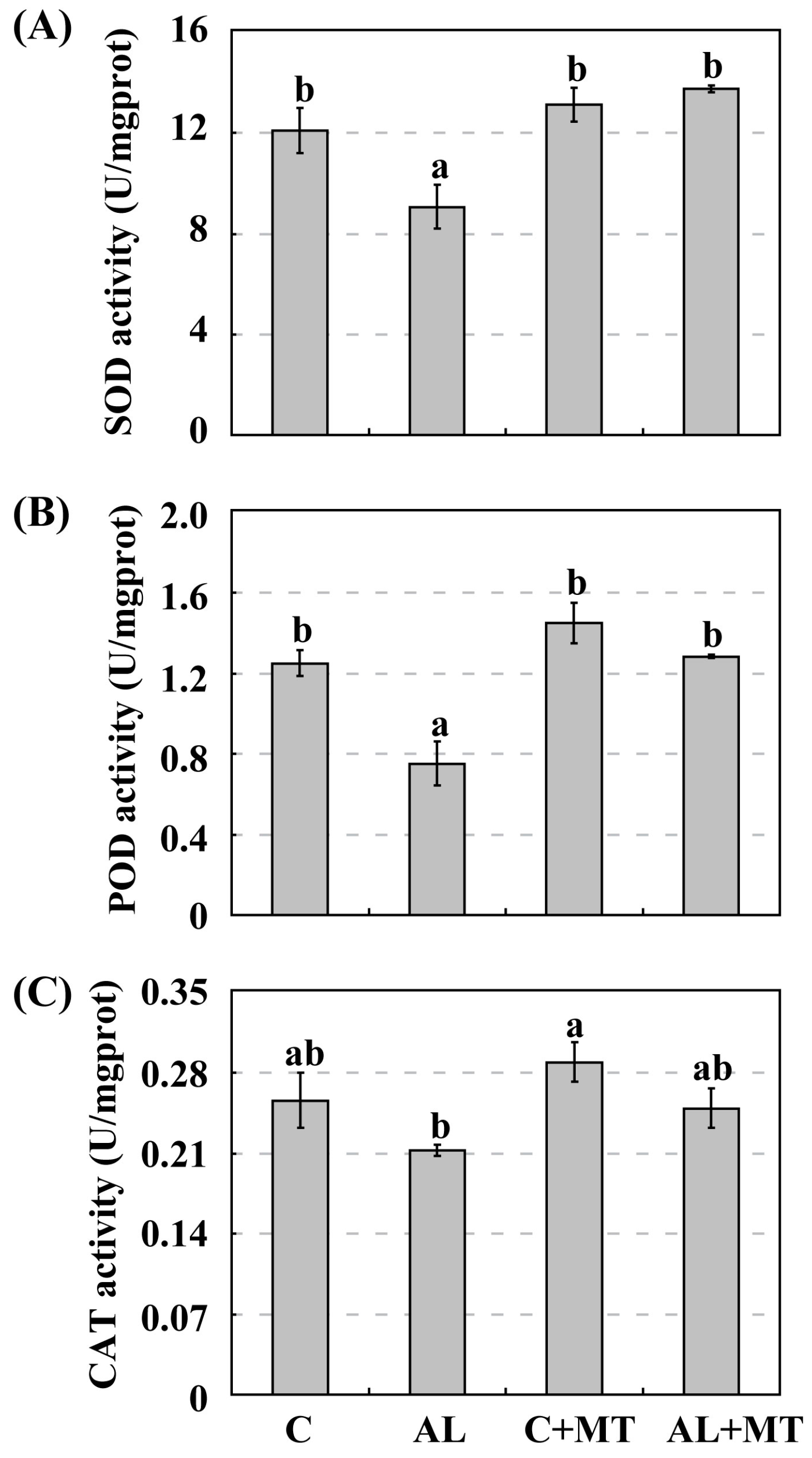
As a versatile antioxidant, melatonin acts to remove excess free radicals, such as ROS and reactive nitrogen species (RNS), from cells when plants are exposed to adverse environments [
34,
50,
51]. We also noted that the induction of alkaline stress was associated with higher production of H
2O
2 and O
2− when compared with the control group, and that lipid peroxidation and membrane damage were more severe under conditions with high pH. However, the application of melatonin reduced MDA concentrations and the accumulation of ROS when stress conditions were introduced, thereby preserving membrane integrity. During the removal of ROS, an antioxidant is converted to an oxidized form that has less scavenging activity. However, melatonin can be regenerated and, similarly, its metabolites can efficiently react with free radicals. Moreover, melatonin boosts the action of other antioxidant systems to remove excess free radicals [
52,
53,
54]. This has been demonstrated in experiments with
Malus sp., where melatonin directly scavenges H
2O
2 and enhances the activities of CAT and another antioxidant enzyme, ascorbate peroxidase, to detoxify H
2O
2 in plants under drought stress [
33]. In watermelon, local application of melatonin activates those enzymes to remove excess ROS and make plants tolerant to cold stress [
55]. Our monitoring of enzyme activity showed that SOD and POD were inhibited under alkaline stress, but melatonin treatment caused those activities to return to nearly the same level as detected under normal conditions. The detected CAT activity was also inhibited moderately by alkaline stress, and melatonin slightly elevated CAT activity. Although alkaline soils can be the source of numerous physiological challenges for plants, melatonin is still an effective broad-spectrum antioxidant that confers stress tolerance to
Malus seedlings.
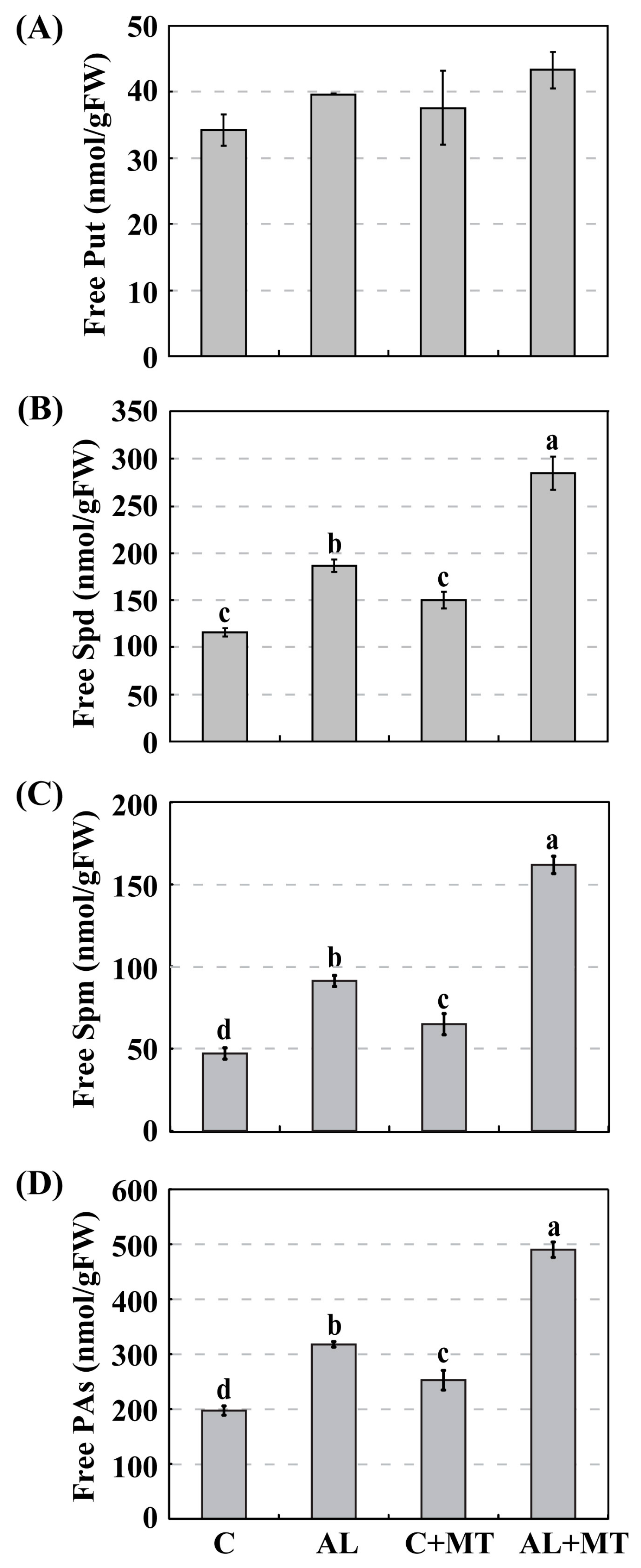
The accumulation of PAs is an integral part of the metabolic strategy by which plants survive under abiotic stresses. This is achieved through either exogenous applications of PAs or the generation of transgenic plants with altered expression of PA-related genes [
56,
57]. Increased PA levels enable plants to respond both spatially and temporally in order to avoid and overcome the effects of stress [
19]. In alkaline-stressed tomato seedlings, Put is dramatically accumulated [
58]. However, we did not find any significant difference in Put accumulations between the normal and stress treatments. Nevertheless, accumulations of Spd and Spm were much higher under the latter experimental condition, implying that the role of PAs in stress responses is both conserved and unique among species. We found it interesting that exogenous melatonin improved the levels of Spd and Spm in stressed seedlings, which suggested the possibility of direct or indirect cross-talk between melatonin and PAs when determining the degree of stress tolerance.
Melatonin might also have a positive role in the PA signaling pathway. For example, exogenous melatonin can significantly increase the accumulation of nitric oxide (NO) to increase plant tolerance to an iron deficiency. However, NO production is inhibited in PA-deficient plants that are sensitive to a lack of iron, which suggests that the positive function of melatonin in plants under Fe-deficient conditions depends upon PA-induced NO production [
59]. The NO signaling pathway is considered a downstream signal in the melatonin signaling pathway [
60]. Meanwhile, these PAs have also been shown to interact with NO in alkaline-stressed plants [
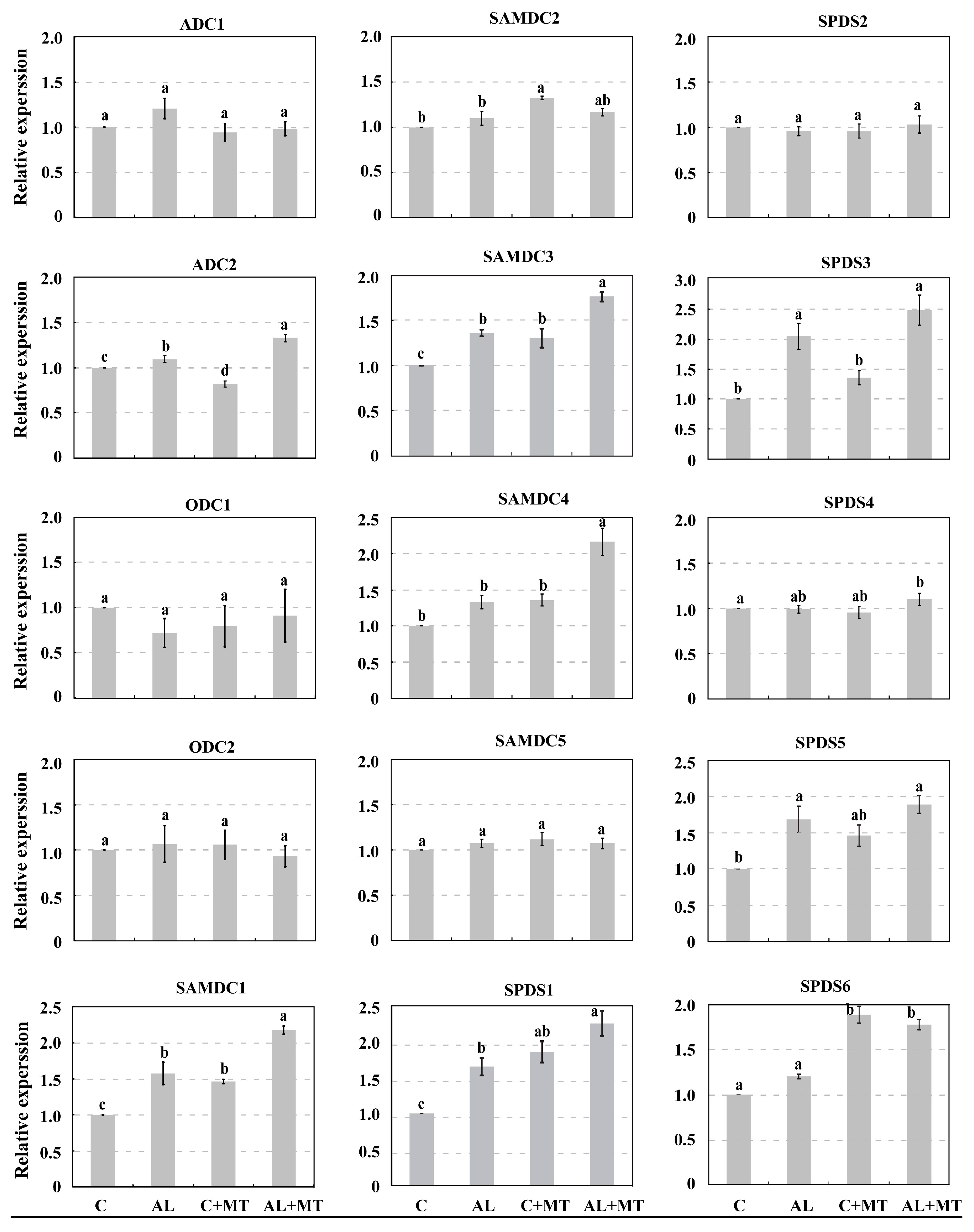
58], implying indirect relation between PAs and melatonin in plants under alkaline stress. In the present study, we found that the contents of PAs were elevated by melatonin application under alkaline stress. Besides, melatonin up-regulated the expression of several genes involved in PA biosynthesis. For melatonin treated seedlings, expressions of
SAMDC1/3/4 and
SPDS1/3/5/6, which are mainly involved in spermidine and spermine biosynthesis, were induced under alkaline stress, leading to increased accumulations of those particular PA, suggesting direct relationship between PAs and melatonin in plants under alkaline stress. Such direct evidence has also been confirmed in plants under low temperature stress. For example, cold-induced apoptosis was inhibited by exogenous melatonin in carrot suspension cells, and notable increases in Put and Spd levels were observed in melatonin-treated cells, which inhibited DNA laddering under cold stress [
61]. In cucumber seedlings, melatonin mediated chilling stress tolerance was also been proved partly via the regulation of PAs metabolism. Melatonin increased the content of Put, Spd, and stabilized Spm under chilling stress [
62]. When
Prunus persica fruits treated with melatonin in cold storage, the expression of
PpADC and
PpODC were elevated, causing an increase in PAs contents [
63]. Therefore, it is speculated that PAs metabolism might directly functions in melatonin signaling pathway to confer tolerance to alkaline stress. PAs may function indirectly in the melatonin signaling pathway through NO. Melatonin, polyamine and NO, they may role in the same network under alkaline stress, which calls for further study.
In summary, we have shown that exogenous melatonin mitigates the adverse effects that alkaline stress can have on the growth of Malus hupehensis seedlings. Treatment with this molecule alleviates damage to cell membranes, reducing lipid peroxidation and ROS accumulations when plants are later exposed to alkaline conditions. Melatonin positively alters RSA, elevates the activity of antioxidant enzymes, and increases levels of polyamines by enhancing the expression of genes involved in their biosynthesis. Further investigations should focus on the mechanism that regulates the relationship between melatonin and PAs.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}