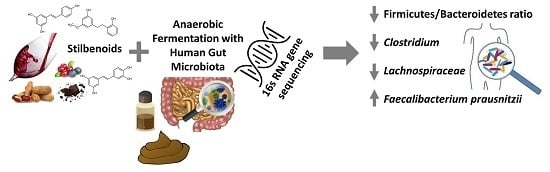
Both parametric and non-parametric statistical tests were used to identify taxa of interest at the phylum, family, and species level based on two comparisons. The statistical tests were used as a tool to identify potential significantly affected taxa, and should not be interpreted as a portrayal of definite statistical significance (for those with p values in the range) due to the small sample size (four donors). The identified taxa reported p < 0.075 for at least one p value (paired sample t-test and/or Wilcoxon signed-rank test) for both comparisons 1 and 2. Comparison 1 used as a baseline the relative abundance of the 24 h control, and compared this value to each of the six stilbenoid fermentations. Comparison 2 used as a baseline the magnitude of change (growth or decline) in relative abundance between Control 0 h and Control 24 h, and compared this value to the magnitude of change between Control 0 h and each of the six stilbenoid fermentations.
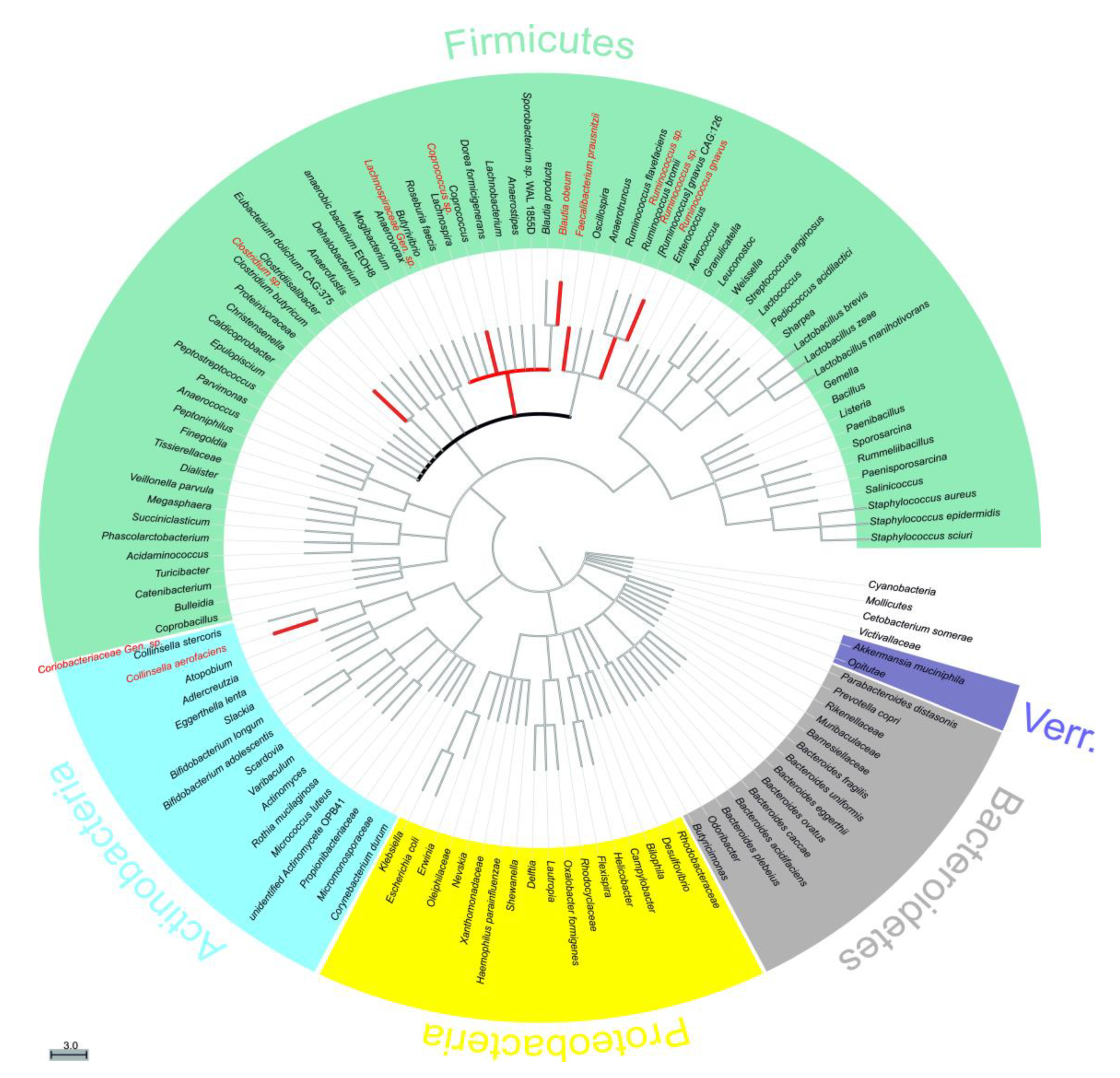
Figure 3 displays these identified taxa in the form of a phylogenetic tree sorted by phylogenetic distance. The corresponding
p values are listed in
Appendix A,
Table A2, and the corresponding relative abundance box plots are shown in
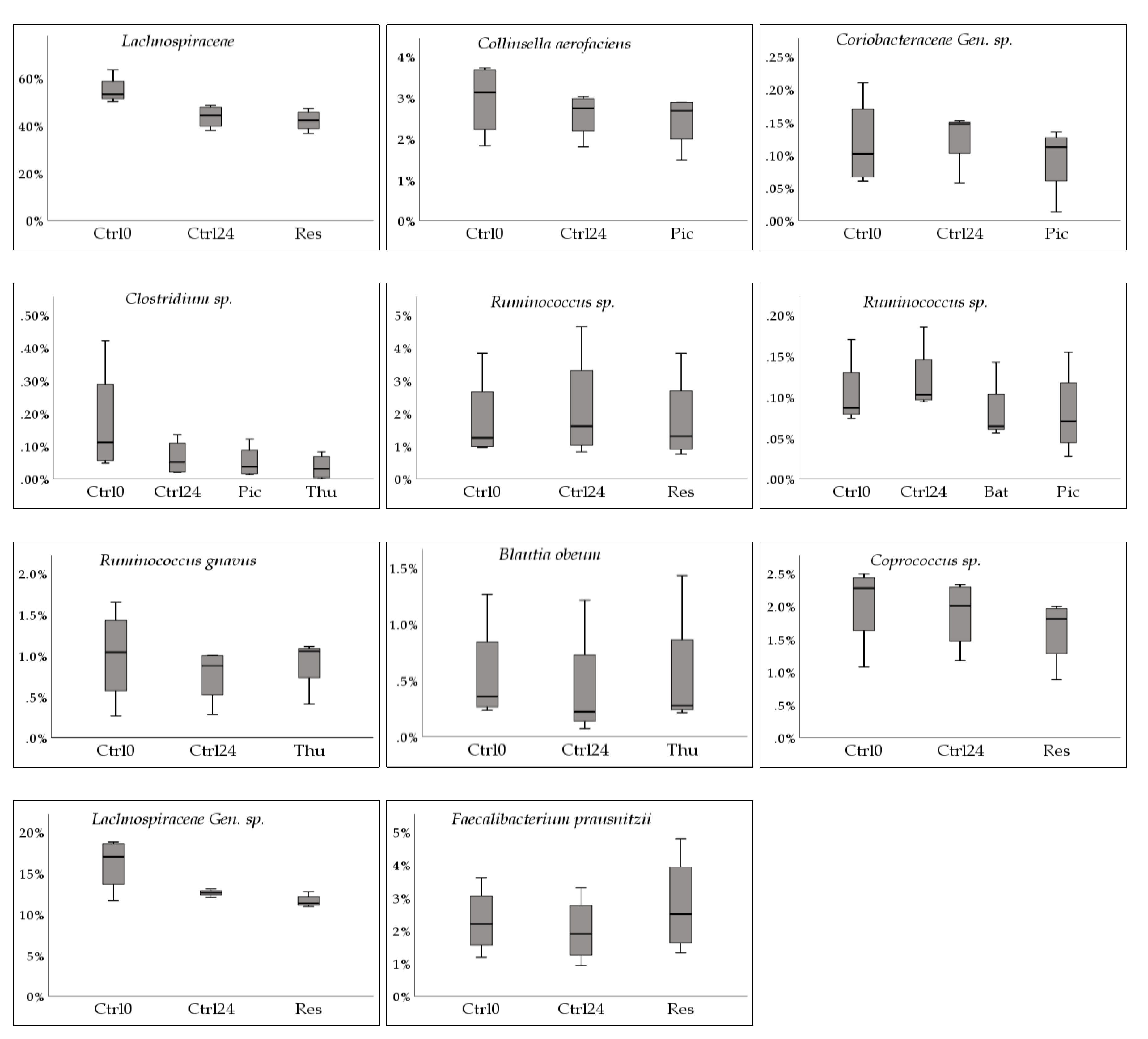
Figure 4. Each comparison (1&2) is shown separately in
Appendix A,
Table A3 and
Table A4, and list additional taxa. Clustered bar graphs of bacterial composition at the phylum and family levels can be seen in
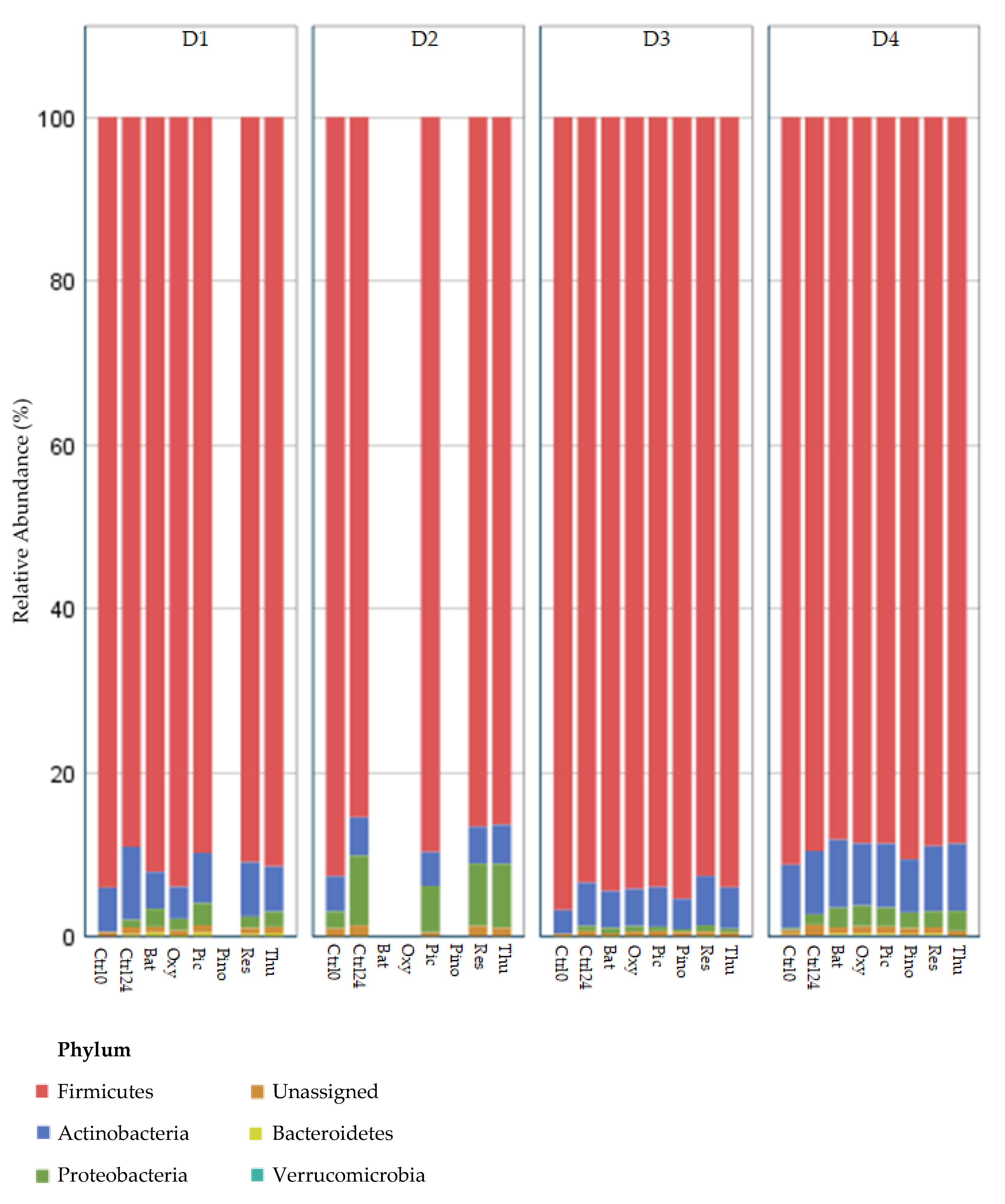
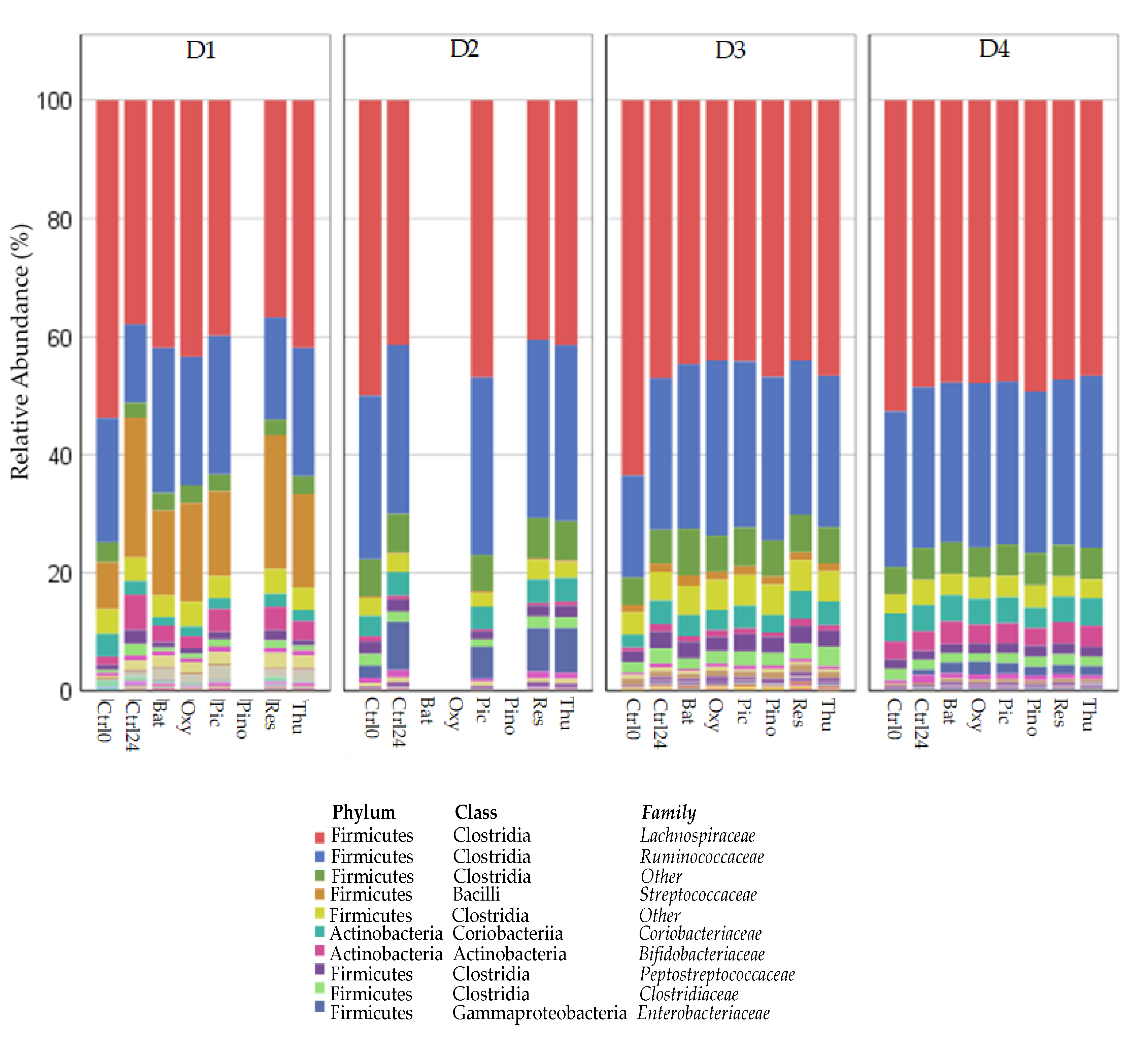
Appendix A,
Figure A1 and
Figure A2.
Table 2 displays how our study compares to findings and observations from other studies regarding the effect of the selected stilbenoids on a specific taxon.
2.3.1. Decrease in Relative Abundance
A decrease in relative abundance was observed for several taxa under some of the tested stilbenoids. The most frequently observed response was a further decrease of the relative abundance of a specific taxon as compared to the 24 h control by either Res, Pic or Thu. For example, for
Clostridium sp. there was a decrease of −54.2% ± 28.8% for Ctrl24, while the decrease caused by Pic and Thu were of a greater magnitude, −62.9% ± 28.0% (t(3) = 3.960, p = 0.029) and −79.3% ± 22.6% (t(3) = 3.901, p = 0.030), respectively. Similar responses were observed, albeit at different magnitudes, for family
Lachnospiraceae, and species
Coprococcus sp.,
Collinsella aerofaciens, and
Lachnospiraceae Gen. sp. At the genus level,
Clostridium decreased under all tested stilbenoids in our study. Previous findings, as listed in
Table 2, observed that several species from the genus
Clostridium, which includes both commensal and deleterious species, had been shown to decrease with resveratrol [
2,
12].
A second observed response was a decrease in relative abundance while the 24 h control increased. This was observed by three species,
Ruminococcus sp. (−3.2% ± 69.1%, t(2) = 4.448, p = 0.047 under Bat; −7.0% ± 69.4%, t(3) = 8.253,p = 0.004 under Pic; −41.1% ± 50.9%, t(3) = 1.953, p = 0.146 under Thu)
, Ruminococcus sp. (−3.3% ± 12.7%, t(3) = 3.947, p = 0.029 under Res), and
Coriobacteriaceae Gen. sp. (−0.9% ± 94.2%, t(2) = 6.272, p = 0.024 under Oxy; −3.7% ± 90.6%, t(3) = 3.261,p = 0.047 under Pic; −39.2% ± 10.0%, t(3) = 1.726, p = 0.183 under Thu), while they increased in the 24 h control (27.8% ± 80.6%, 32.2% ± 68.5%, 15.5% ± 20.8%, respectively). Regarding
Ruminococcus, this may not be a favorable response according to recent research that points to a high proportion of long-chain dietary fibers degraders, butyrate producing bacteria such as
Ruminococcus, Eubacterium, and Bifidobacterium as being part of healthy gut microbiota [
11,
40,
41,
42]. The
Ruminococcus genus has previously been identified as one of the three taxa, besides
Bacteroides and
Prevotella, that define the enterotype concept, which could help in explaining variability in responders/non-responders in intervention studies [
43]. In regards to
Coriobacteriaceae, it has been noted that many species that metabolize phenolics belong to this family, however, its potential health implications are still poorly understood [
6]. Nevertheless, one important aspect of this family is that all identified
S-equol-producing bacteria, except for the genus
Lactococcus, belong to it [
44,
45].
A third observed response was a decrease in relative abundance while the 24 h control also decreased, but with a larger magnitude. This was observed for
Blautia obeum, which was recently reclassified, its former name being
Ruminococcus obeum [
46].
Blautia has been considered one of the major representatives of the Firmicutes phylum due to its relatively high abundance [
15]. This species experienced a decrease in relative abundance by thunalbene (−5.6% ± 32.1%, (t(3) = 3.763, p = 0.033), but at a lower magnitude than the control at 24 h (−29.8% ± 35.6%). A decrease of
Blautia, at the genus level, was also reported in a study conducted on mice fed a phenolic-enriched tomato diet, as well as in a study of human fecal fermentation study after consumption of phenolics from tart cherries [
26,
31]. These findings, along with our study, suggest that certain phenolics may cause a decrease in this genus, but at a lesser magnitude than without it. This taxon also appears to be a butyrate-producing microbe whose reduction has been correlated with decreased production of butyrate [
47].
Eight of the identified taxa belonged to the family
Lachnospiraceae. There was no consistent response from the tested stilbenoids within this family however, the most frequent response was a decrease in relative abundance. This decrease was also observed in a study where rats were supplemented with the stilbenoid pterostilbene in their diet. In that study,
Lachnospiraceae was significantly reduced in each tested group when compared to baseline levels [
33].
2.3.2. Increase in Relative Abundance
An increase in relative abundance with no change in the 24 h control was observed for
Faecalibacterium prausnitzii under Res (36.6% ± 88.0%, t(3) = −2.806, p = 0.068 under Res), 24 h control (−0.5% ± 62.5%). This species has been previously identified as a butyrate producing bacterium and is regarded as being beneficial. Butyrate production appears to be key in maintaining the colonic epithelium by inducing proliferation of healthy colonocytes. Fiber-poor diets, such as the one our donors were subject to prior to sample donation, have been associated with low butyrate production. One study showed a strong positive correlation between the proportion of
F. prausnitzii and that of butyrate in individuals on a normal diet, and the reduction in
F. prausnitzii on switching to a fiber-free or fiber-supplemented diet correlated with the reduction in fecal butyrate [
47,
48]. The gut epithelium is the main body site for butyrate sequestration, and low butyrate production has been connected to inflammatory diseases such as ulcerative colitis [
39,
49]. Unlike acetate producing bacteria, which are widely distributed, there appear to be fewer butyrate producing bacteria such as
S. prausnitzii, E. rectale, E. hallii, and
R. bromii [
38]. It was observed to increase in plant-based, fiber-rich, diets, thus, stilbenoids being phytochemicals, were expected to increase their abundance. Our findings support this with resveratrol.
An increase in relative abundance with a decrease in the 24 h control was observed for
Ruminococcus gnavus under Thu (8.2% ± 40.6%, t(3) = −2.244, p = 0.111 under Thu), 24 h control (−12.9% ± 30.7%). The observed p value, along with the box plot in
Figure 4, show that
R. gnavus’ increase was not as pronounced as that of
F. prausnitzii. Both of these taxa tend to be quite reduced in inflammatory bowel diseases such as Crohn’s disease [
50,
51].
Although it was detected in only one of our donors,
Akkermansia muciniphila was observed to be enhanced by resveratrol. This species has been previously observed to be enhanced by pterostilbene, which has shown to exhibit similar cellular effects to resveratrol. One of these is that both phenolics have been hypothesized to mimic caloric restriction effects at the molecular level, thus modifying the gut microbiota, especially enhancing
A. muciniphila [
33].
These findings emphasize the importance of trying to get to the lowest possible taxonomic level to better characterize the gut microbiota. As can be seen from our study, species within the same family level are not all uniform in their responses. Higher taxonomic levels are quite useful, and can make experiments and data processing much more manageable; however, care must be taken in generalizing for every member of a taxon.
Whether the microbiota response is a decrease or an increase in relative abundance, effects are more frequently attributed to resveratrol and piceatannol, followed by thunalbene and batatasin III. This difference may be related to their chemical moieties. All stilbenoids share a basic C6-C2-C6 structure, differing only in the presence or absence of a C-C double bond on -C2-, and on the type and position of functional groups, mainly hydroxyl (-OH) and o-methoxyl (-OCH
3) groups on the aromatic rings. In phenolics, -OH groups play an important role on their bioactivity, and their substitution by -OCH
3 groups has been shown to reduce their bioactivity [
52,
53,
54]. -OH groups are good hydrogen donors, are considered very reactive and potent radical scavengers, are key in the general antioxidant mechanism of resveratrol, and it has been shown that phenolics with more -OH groups exhibit higher capacity for enzyme inhibition than those with -OCH
3 groups [
53,
54,
55,
56,
57]. Enzyme inhibition capacity has also been shown to be affected by hydrogenation of the C-C double bond on -C2-, which decreased enzyme inhibition [
54,
58,
59,
60]. This suggests that phenolics with -OH moieties and C-C double bond on -C2- may be more bioactive than those with -OCH
3 moieties and lacking a C-C double bond on -C2-. Resveratrol and Piceatannol have three and four -OH groups respectively, as well as a C-C double-bond on -C2-. They were the two stilbenoids that were most frequently attributed effects on the GM in this study. These were followed by thunalbene, which is O-methylated and has a C-C double bond on -C2-, and by batatasin III, which is O-methylated and lacks a C-C double bond on -C2-. Regarding demethylation, a recent study reported a demethylated colonic metabolite of the phenolic curcumin by
Blautia sp. MRG-PMF1 [
61]. Thunalbene is O-methylated and, as reported earlier,
Blautia sp. experienced a decrease in relative abundance under thunalbene, but at a lower magnitude than that of the control. Regarding C-C double bond reduction, Bode et al. showed that
Slackia equalifaciens and
Adlercreutzia equolifaciens were able to metabolize resveratrol to dihydroresveratrol by reduction of the C-C double bond, but could not identify any bacteria for the -OH cleavage that produced two other metabolites [
8]. Reduction of the C-C double bond by GM has also been shown for other phenolics such as isoflavones and hydroxycinnamates, while -OH cleavage for lignans and phenolic acids [
19,
62,
63,
64]. How chemical moieties affect metabolite production by microbial strains and bioactivities such as antioxidant activity, enzyme inhibition, quorum sensing, and others is outside the scope of our study; nevertheless, it’s an important avenue for ongoing and future research.
The interpretation of the results from GM studies such as this one should take into consideration the concept of inter-individual variability. This concept is well known in the literature, the most well-known example being the difference between individuals whose microbiota are either producers or non-producers of the
S-equol phytoestrogen. Oral administration of
S-equol results in improvement of certain cardiovascular disease biomarkers, but only on those who are producers [
20,
47]. Although our sample size is small, differences among donor GM composition can be visualized in
Figure A1 and
Figure A2. Donor D2, for example, appears to have a very atypical microbial composition when compared to the other three donors.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





