1. Introduction
Pyropia is an important genus of red algae. According to the Food and Agriculture Organization of the United Nations (FAO), the laver production was about 1.8 million tons (fresh weight) in 2016, with a commercial value of more than USD 1.5 billion [
1]. In China, naturally there are about 22
Pyropia species, among which the two most important species are
P. yezoensis and
P. haitanensis [
2].
P. haitanensis is cultivated on a large scale and shows the highest yield among all laver species [
1]. It is extensively distributed in the coastal areas of Guangdong, Fujian, Zhejiang, and Jiangsu provinces [
3]. At present, the annual output of
P. haitanensis is about 88,000 tons (dry weight), accounting for 75% of the total output of nori in China and more than 50% of the total output of nori in the world [
1]. It is a nutrient-rich marine food material and a pillar product of the rural economy of China.
The nutritional value and flavor are closely related to the content of protein and amino acids.
Pyropia is favored by people due to its characteristic taste, which results from relatively large amounts of flavor amino acids, such as glutamic acid, aspartic acid, and glycine [
4]. At present, the research on flavor amino acids in
Pyropia is mainly limited to the comparison of nutrient contents [
5,
6], the extraction methods [
7], the taste index of food [
8,
9], and the factors affecting the content of amino acids. The content of flavor amino acids is mainly related to external nutrition supply and harvesting time [
6,
9,
10]. Li et al. [
10] reported that the amino acid contents could increase under the combined application of nitrogen and phosphorus fertilizers in
P. yezoensis. However, few studies have focused on the functional characterization of key enzymes in the amino acid metabolism in red algae.
Due to its high content, glutamic acid is considered to be the main umami amino acid in
Pyropia [
6]. There are two ways to synthesize glutamic acid in organisms, one is dependent on glutamine synthetase (GS)/glutamate synthase (GOGAT), and the other is dependent on glutamate dehydrogenase (GDH) [
11]. Although GS/GOGAT is considered to be the main pathway of ammonium assimilation, GDH can be used as an auxiliary approach and play an irreplaceable role in carbon and nitrogen metabolism in organisms.
The biosynthetic pathway of glutamic acid dependent on GDHs is as follows (
Figure S1): glucose is converted to pyruvate through the Embden-Meyerhof-Parnas (EMP) pathway; the latter is converted to α-oxoglutarate in the tricarboxylic acid (TCA) cycle; α-oxoglutarate is catalyzed to glutamic acid by glutamate dehydrogenase (GDH) [
11]. GDH is the important enzyme in glutamic acid biosynthesis, and is also one of the core enzymes in nitrogen metabolism [
12]. GDHs can be divided into four distinct classes, of which GDH-1 and GDH-2 are small hexamer enzymes that are widely distributed in animal and plant tissues and play an important role in the process of ammonia assimilation [
13]. GDH-3 has a large molecular weight, which acts on the catabolism of glutamate. GDH-4, discovered in eubacteria, is about 180 kDa in size and has NAD
+ specificity [
13]. According to different types of coenzymes, GDHs can be divided into the following three types: the first type uses NAD(H) as a coenzyme (EC 1.4.1.2), the second type is GDH that relies on NADP(H) as a coenzyme (EC 1.4.1.4), and the third type GDH (EC 1.4.1.3) can rely on NAD(H) or NADP(H) as a coenzyme [
14]. In higher plants, GDH-catalyzed reaction is not considered to be the main pathway for glutamic acid biosynthesis, because GDHs have a relatively high
Km value for ammonium ions. In contrast, GDHs in some lower organisms exhibit higher affinity for ammonium and play a more significant role in ammonium assimilation [
15]. GDHs can catalyze the synthesis of glutamic acid from ammonia and α-oxoglutarate, which serve as a link between carbohydrate and amino acid metabolism [
16,
17]. Moreover, they play a key role in controlling glutamic acid homeostasis and supplementing the lack of carbon under certain physiological conditions [
14]. GDHs also appear to be more active when energy supply is low due to abiotic stress [
18,
19]. They have been functionally verified in vitro in many organisms, such as
Salmonella typhimurium and
Rumen ciliate [
20,
21]. In addition, the function of GDHs has also been verified by in vivo experiments in higher plants but not in algae. The content of glutamic acid in tomato fruit transformed with the
Aspergillus GDH gene was twice that of the control group [
22]. However, the function of GDHs in
Pyropia has not been investigated, although it is closely related to the quality traits of laver.
In this study, we cloned two GDH genes from the red alga P. haitanensis, investigated their sequence structures and enzymatic characteristics, and examined their transcription profiles under various abiotic stresses. We aim to verify the role of GDHs in the biosynthesis of glutamic acid and to provide a reference for the improvement of quality traits of P. haitanensis.
3. Discussion
Glutamic acid is an important flavor substance, but its metabolic pathways and relevant catalytic enzymes in red algae are scarcely studied. In this study, we measured the content of glutamic acid in
P. haitanesis sampled from four different locations of China and found that the content of glutamic acid was higher in
P. haitanesis from the southern region (Putian) than in that from the northern region (Yancheng). Moreover, the correlation analysis of glutamic acid content and the expression of PhGDHs showed a consistent trend, indicating that PhGDHs might be related to glutamic acid metabolism. In higher plants, the GS/GOGAT is considered to be the main pathway of ammonium assimilation. However, our unpublished data on the RNA-seq result of
P. haitanensis samples collected from different harvesting stages showed that GS unigenes were found but with very low RPKM (Reads Per Kilobase per Million mapped reads) values (<0.5) (
Table S2). This might imply the lower activity of GS in
P. haitanensis. Therefore, we conjected that the PhGDHs could participate in the glutamic acid biosynthetic pathway. We further identified two
GDH genes from
P. haitanensis,
PhGDH1 and
PhGDH2. They have similar domains to other GDHs from red algae, which shows that they do have the function of dehydrogenase. We compared their sequence characteristics as well as in vitro enzyme activities and aim to elucidate possible mechanisms for the flavor and stress resistance ability of
P. haitanensis.
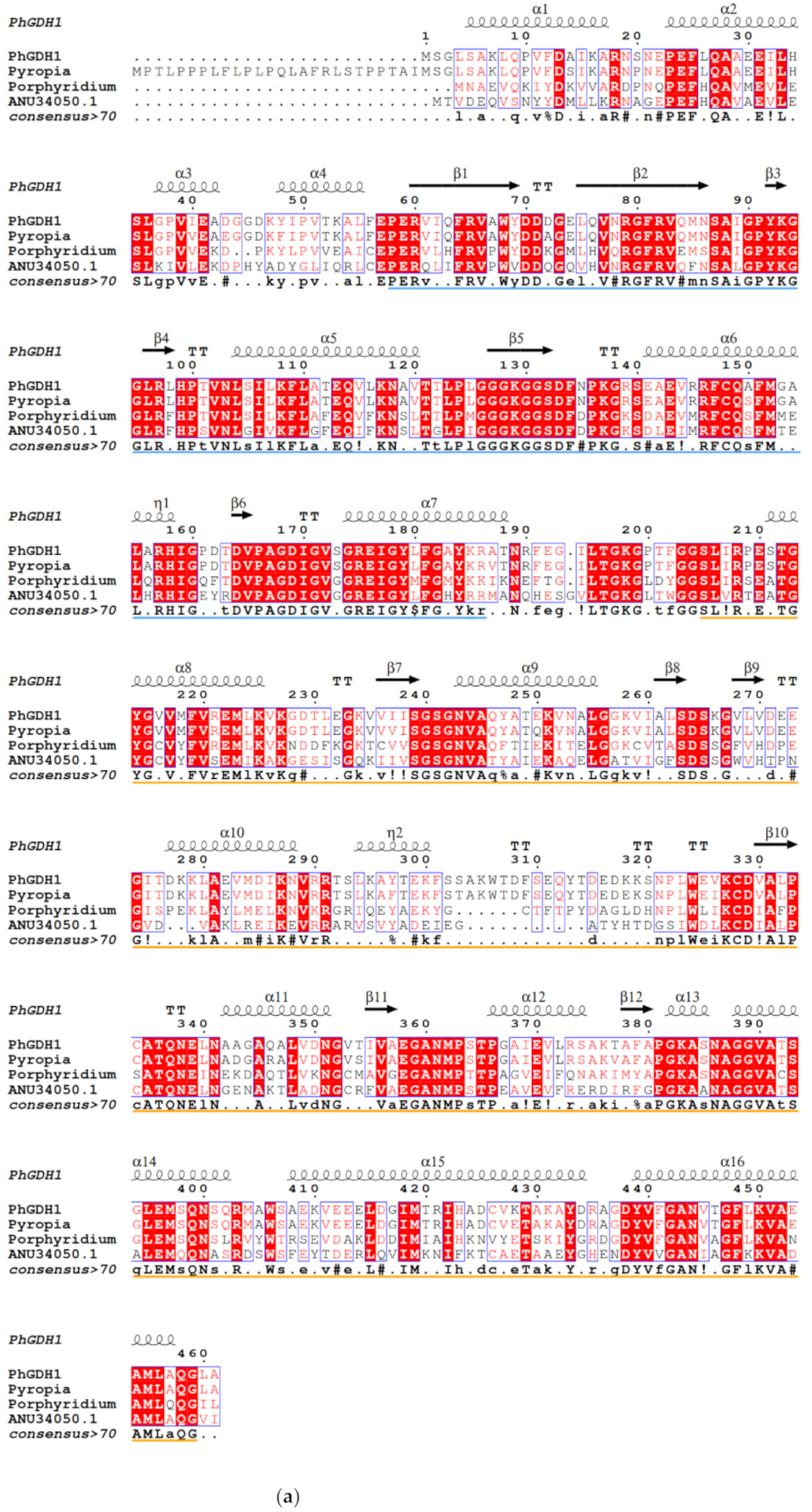
GDHs can be divided into four categories according to their metabolic specificity and subunit size [
23], GDH-1 and GDH-2 are small hexamer enzymes, while GDH-3 and GDH-4 have a large molecular weight. In this study, both PhGDH1 and PhGDH2 are small hexameric enzymes (~50 kDa), which belong to GDH-1 or GDH-2. Generally, in hexameric GDHs, each subunit is divided into two domains, and there is a deep cleft between the two domains [
24]. Domain I is mainly composed of the N-terminus of the polypeptide chain, responsible for the symmetrical binding of subunits, and participates in the formation of hexamers. Domain Ⅱ is composed of the C-terminal part of the chain and participates in the binding of the cofactor [
24]. In PhGDHs, each subunit can also be divided into two domains. According to the secondary structure prediction results, both contain classic Rossmann fold for binding NAD(P)H. Both PhGDH1 and PhGDH2 can use NADH or NADPH as coenzymes, so they may belong to the third type GDH (EC 1.4.1.3). However, they show much higher activity against NADH than that for NADPH, so NADH is the main cofactor for PhGDH1 and PhGDH2. To confirm the involvement of candidate residues in the binding of NADH in
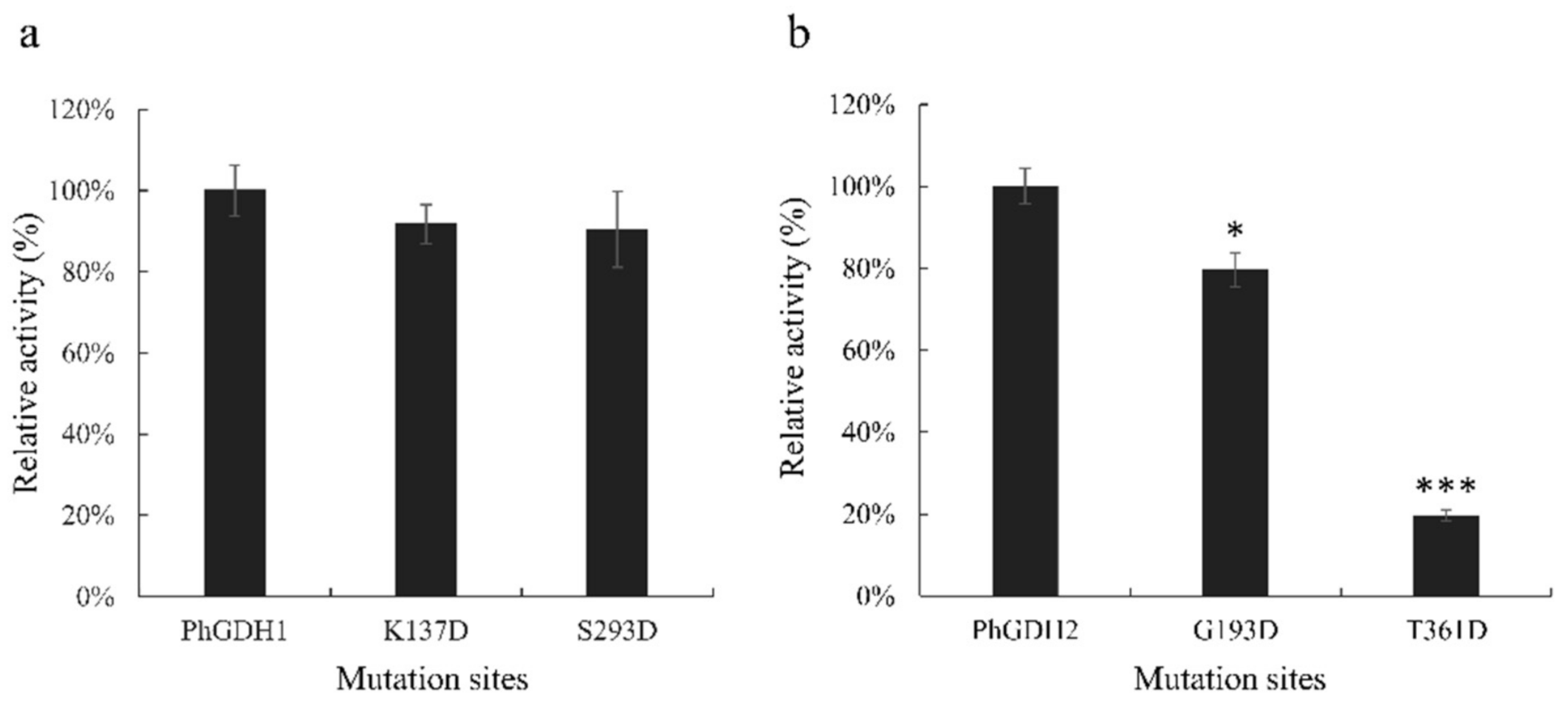
P. haitanensis, we mutated the putative residues Lys
137 and Ser
293 of PhGDH1, and Gly
193 and Thr
361 of PhGDH2 to aspartic acid. These residues in the same position in the GDH from
Corynebacterium glutamicium have been confirmed to be active sites [
24]. All the mutated genes can express soluble proteins in
E. coli, suggesting that none of these sites prevented the protein from folding efficiently. The activities of K137D and S293D decreased slightly; however, the G193D and T361D activities significantly decreased, which indicates that Gly
193 and Thr
361 are important for the binding of NADH in
P. haitanensis. Notably, these two sites are different in GDHs from
Gracilariopsis chorda and
Galdieria sulphuraria (
Figure 1), suggesting Gly
193 and Thr
361 may be novel NADH-binding sites in
P. haitanensis.
GDHs catalyze a reversible reaction. We therefore tested the reaction rate in the two directions in vitro. The reaction rate in the direction of glutamic acid degradation was much lower (
p < 0.05), implying the predominant role of PhGDHs catalyzing the biosynthesis of glutamic acid. In the ammonium assimilation direction, PhGDH1 and PhGDH2 had similar optimal reaction temperature and pH. Both PhGDHs exhibited the highest catalytic efficiency at 25 °C, which was close to the suitable growth temperature of
P. haitanensis (20 °C). Their optimal reaction temperature is close to the growth temperature of
Laccaria bicolor (30 °C) [
25] and
Bacillus subtilis natto (30 °C) [
26], but lower than that of
Phormidium laminosum (60 °C) [
27] and
Pyrococcus horikoshii (90 °C) [
28]. We speculate that the optimal reaction temperature of GDHs may be related to the growth temperature specific to different organisms. The two PhGDHs are suitable to catalyze the reaction in an alkaline environment (the optimal pH values of PhGDH1 and PhGDH2 are 8.0 and 8.5, respectively), which may be related to the weak alkalinity of seawater. However, PhGDH2 is more sensitive to acidity than PhGDH1, and PhGDH2 lost most of its activity at pH 6.5. It has been previously reported that the optimal pH values for the catalytic reaction of GDHs from
Bryopsis maxima [
29],
Pyrococcus horikoshii [
28], and
Gigantocotyle explanatum [
30] are 7.5, 7.6, and 8.0, respectively. Although these GDHs possess different optimal pH values, they all exhibit higher catalytic activities in the alkaline environment.
For the three substrates, the
Kcat values of PhGDH1 are much higher, which means it has higher catalytic rate. Both PhGDHs had similar
Km values (0.16 mM and 0.104 mM) for α-oxoglutarate, which are lower than those of GDHs from
Pyrococcus horikoshii (K
m = 0.53 mM) [
28] and
Thermus thermophilus (
Km = 3.5 mM) [
31]. However, PhGDH2 showed a much lower
Km value for NADH compared to PhGDH1, which may be due to certain differences in the cofactor-binding sites between the two enzymes. The
Km value for NH
4+ can reflect the ability of ammonia assimilation, and the
Km values of PhGDH1 and PhGDH2 for (NH
4)
2SO
4 are remarkably lower than that of GDHs in
Cucurbita pepo (
Km = 33.3 mM) for NH
4+ [
32]. PhGDH1 and PhGDH2 present much higher affinity for NH
4+ than GDHs from most higher plants (
Km = 10–80 mM) [
33]. It is reasonable to speculate that they can assimilate ammonium more effectively. This phenomenon may be related to the growing environment of
P. haitanensis, where it needs to adapt to periodic dehydration stress and consequent high-temperature stress, while the habitat of the higher plants is more stable. When under abiotic stresses, the concentration of ammonia in
P. haitanensis cells increases. Therefore, PhGDHs with higher affinity for NH
4+ is vital for the survival of
P. haitanensis. Overall, the
Km values of PhGDH2 for the three substrates were lower compared to PhGDH1, especially that for the NADH, but its
Kcat/
Km values were also lower. Therefore, PhGDH1 has higher catalytic efficiency than PhGDH2, while PhGDH2 has a higher affinity for the three substrates.
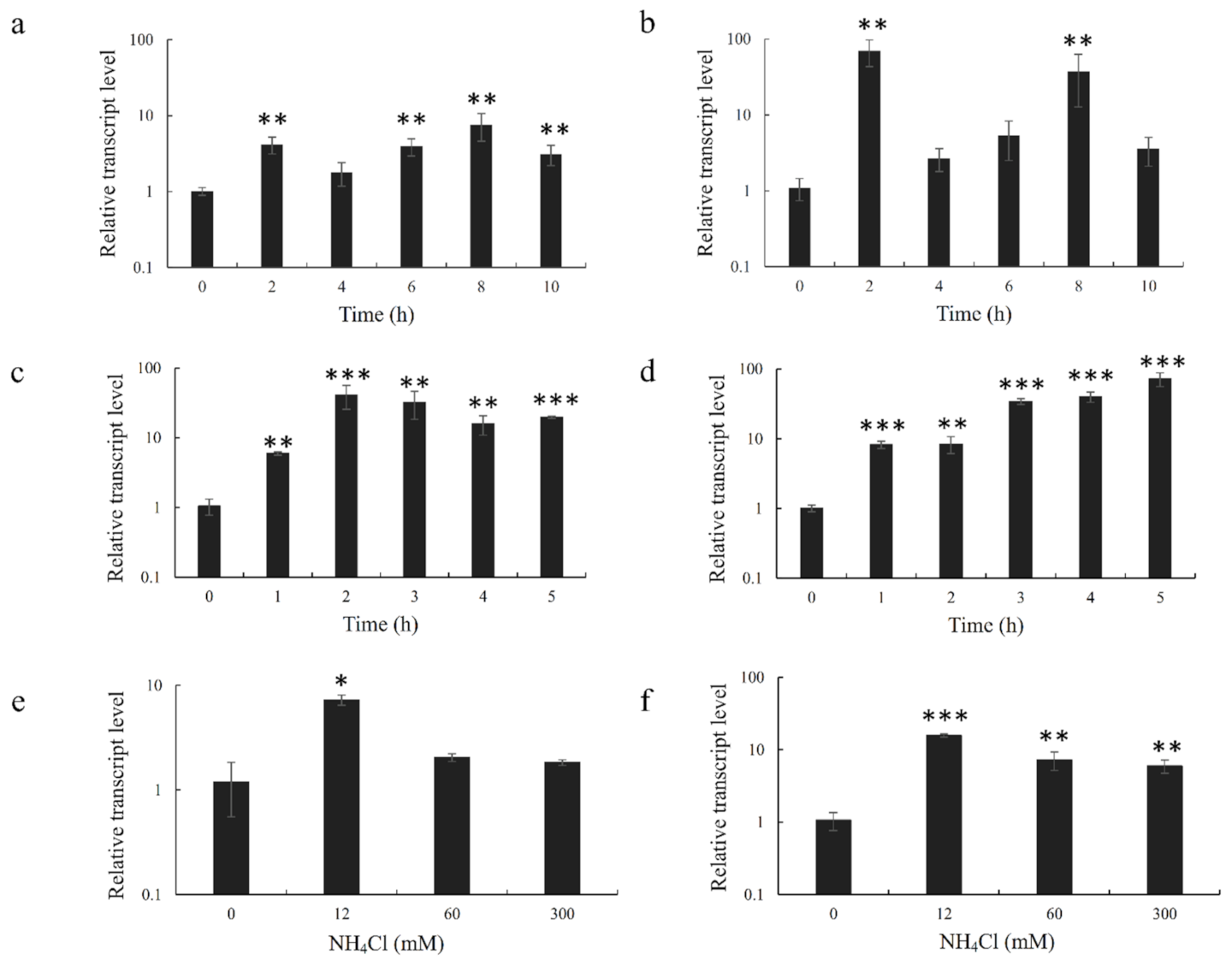
P. haitanensis inhabits the intertidal zone and must undergo periodic dehydration and rehydration processes. Therefore, it has evolved a set of molecular mechanisms to cope with abiotic stresses. Many studies have reported that GDHs are involved in plant response to abiotic stresses. For example, Mena-Petite et al. [
34] showed that GDH activities increased by more than 170% when
Pinus radiata seedlings were subjected to drought stress. In addition, Tang et al. [
35] observed that the GDH activity increased under heat stress in coral. In the present study, we found that abiotic stresses can significantly increase the expression levels of both PHGDH1 and PHGDH2 (
p < 0.05). At the beginning of the abiotic stress, the expression of the two
GDH genes rose rapidly and maintained at a high level throughout the stress compared with the control group. As the stress intensity increased, their expression levels declined, which may be due to the damage of the algal cells caused by the prolonged abiotic stress. High temperature induced the most drastic changes in the expression of
PHGDH1 and
PHGDH2. One of the possible reasons is that the activity of glutamate synthase decreases under high-temperature stress, which further aggravates the effect of elevated ammonium ion concentration [
36]. Desiccation or high-temperature stress leads to the hydrolyzation of a large number of proteins, resulting in the increase of ammonia in algal cells. If the accumulated ammonia is not scavenged in time, it would have a toxic effect on algal cells [
37]. GDHs can reduce the toxic damage resulted from excessive accumulation of NH
4+ by assimilating α-oxoglutarate and NH
4+ into glutamic acid during carbon metabolism. In addition, glutamic acid can continue to synthesize proline and improve the resistance of plants to abiotic stresses [
38]. In this study, both PhGDHs responded to ammonium salt stress, which provides the evidence that these enzymes can synthesize glutamic acid to resist high-NH
4+ stress. According to the performance of
PhGDH1 and
PhGDH2 under various abiotic stresses, we presume that they play a role to help
P. haitanensis in adapting to the harsh environment of the intertidal areas.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}