1. Introduction
The flavonoid biosynthetic pathway has been well investigated and is thought to be a gold mine for metabolic engineering [
1]. The application of biological engineering of flavonoids is prevalent, and there have been many studies on the modification of color, the change in medicinal plant composition, and the improvement of crop nutrition. The flavonoid class of secondary metabolites mainly contains anthocyanins, flavonols, flavones and proanthocyanidins [
2,
3]. Anthocyanin is essential to the physiological and biochemical activities of plants, including antioxidation, protection of plant damage, stress responses, and attracting animals to pollinate and spread seeds [
4]. In recent years, more and more attention has been paid to anthocyanins because of their prominent role in medical resources [
5].
Flavonoid-related genes are primarily responsible for flavonoid biosynthesis, and the activities of different biosynthesis enzymes control the flavonoid profile [
6]. Flavonoid biosynthesis starts with the general phenylpropanoid pathway, producing 4-coumaroyl-CoA by phenylalanine ammonia lyase (PAL) and cinnamate-4-hydroxylase (C4H). 4-coumaroyl-CoA as precursor enters the flavonoid biosynthesis pathway, leading to different metabolic branches, including the products of chalcones, aurones, isoflavonoids, flavones, flavonols, flavandiols, anthocyanins [
7]. This first committed step in flavonoid biosynthesis is catalyzed by chalcone synthase (CHS), which uses malonyl CoA and 4-coumaroyl CoA as substrates [
8]. The product naringenin chalcone is subsequently catalyzed by chalcone isomerase (CHI), and the product naringenin is a general precursor of flavonols, anthocyanins, proanthocyanidins, flavones, and isoflavones [
9]. Favone synthase I (FNS I) or flavone synthase II (FNS II) participate in the synthesis of flavones from naringenin. Isoflavone biosynthesis starts from naringenin or liquirtigenin, in a reaction catalyzed by Isoflavone synthase (IFS), as the first step [
10]. Naringenin is also converted to dihydroflavonols by flavanone 3-hydroxylase (F3H). The dihydroflavonols are precursors of flavonols and anthocyanins. Flavonol synthase (
FLS) is responsible for the synthesis of flavonols and dihydroflavonol reductase (
DFR), flavonoid 3′-hydroxylase(F3′H), anthocyanidin synthase (ANS), and UDP-glucose: flavonoid 3-glucosyltransferase (UFGT) are responsible for the subsequent biosynthesis of anthocyanins [
10].
The flavonoid pathway-related enzymes F3H, FLS and ANS all belong to the 2-oxoglutarate-dependent dioxygenase (2-ODD) subfamily [
11,
12]. F3H is a key enzyme in directing carbon flow towards the biosynthesis of 3-hydroxylated flavonoids and is responsible for the biosynthesis of flavonols and anthocyanidins [
13]. Down-regulation of
F3H in strawberry resulted in a great decrease in anthocyanin and a moderate decrease in flavonol content [
14]. Overexpression of
Lycium chinense LcF3H in tobacco showed increased content of flavan-3-ols and increased tolerance to drought stress [
15]. However, overexpression of
Sorghum bicolor SbF3H resulted in a subtle impact on flavonol production in tissues with endogenous F3H activities [
13].
Carthamus tinctorius CtF3H showed diametrically opposite expression patterns in different phenotypes with orange-yellow flowers and white flowers when exposed to external methyl jasmonate (MeJA) treatment, which has been identified as an elicitor of flavonoid metabolites [
16]. The above facts suggest that the function of F3H in flavonoid biosynthesis is highly complex, and that its impact on flavonoid profile might be species-specific or phenotype-specific.
Mulberry (
Morus spp., Moraceae) plants are distributed widely in China and East Asia and are known as important economic plants with nutritional, medicinal, and ecological value [
17,
18]. Mulberry fruits are a nutritional foodstuff and are rich in flavonoids and anthocyanins. They are recommended as having antioxidant, antimicrobial, and anti-inflammatory properties [
18]. Previous studies have reported that expression levels of genes involved in anthocyanin biosynthesis, including
CHS1,
CHI,
F3H1,
F3′H1, and
ANS showed a close correlation with anthocyanin content during the fruit ripening process [
19,
20]. Given the complex roles played by
F3H in flavonoid and anthocyanin biosynthesis, it is still necessary to further elucidate how
F3H functions in the flavonoid biosynthesis pathway and anthocyanin accumulation during fruit ripening in mulberry. In the present study, we functionally characterized
F3H in mulberry and found its variety-specific impact on anthocyanin accumulation. The expression patterns of
F3H differ greatly among different mulberry varieties during the fruit ripening process. Generally, down-regulation of
F3H in mulberry can result in a decrease in anthocyanins and a subtle increase in flavonoid content, while overexpression of mulberry
F3H in tobacco also resulted in a significant decrease in anthocyanins. This suggests that more complex regulation mechanisms of flavonoid profile partitioning may possibly exist in mulberry.
3. Discussion
Many previous studies have reported the relationships between structure genes involved in flavonoid biosynthesis and the accumulation of flavonoids and anthocyanins [
19,
23,
24]. In mulberry,
CHS1,
CHI,
F3H1,
F3′H1 (flavonoid 3′-hydroxylase), and
ANS have been reported to correlate with anthocyanin biosynthesis during the fruit ripening process [
19]. Our previous study on
MmCHI1 and
MmCHI2 from
Morus multicaulis also showed that the dominant
CHI MaCHI2 had a positive correlation with anthocyanin accumulation in fruit [
25]. The present study shows that
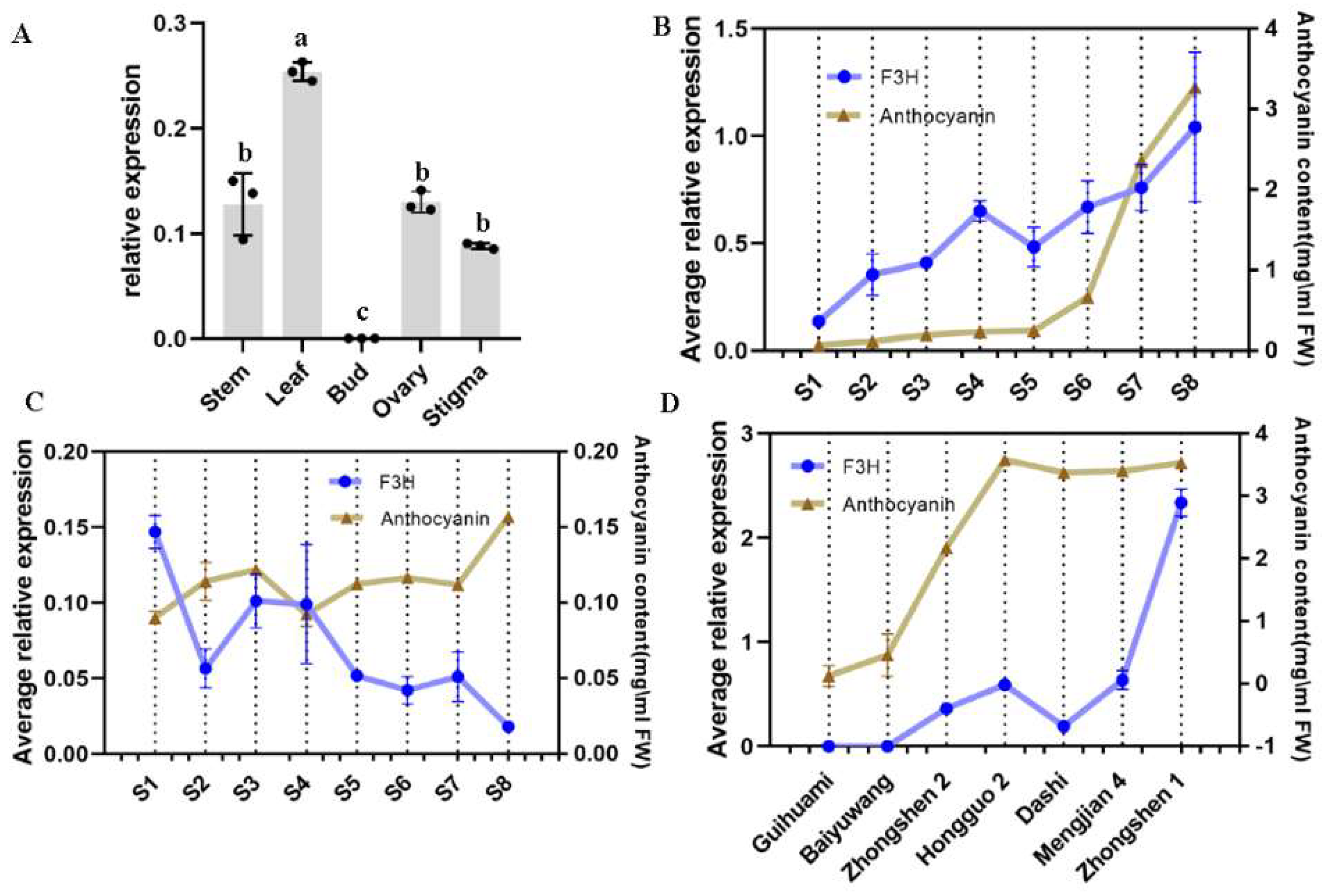
F3H expression levels have a positive correlation with anthocyanin accumulation in anthocyanin-rich mulberry fruits, while a negative correlation was found with anthocyanin content in mulberry fruits that did not require anthocyanin accumulation during fruit ripening process. These results indicate that different mechanisms may exist for regulation of flavonoid or anthocyanin biosynthesis in different mulberry varieties.
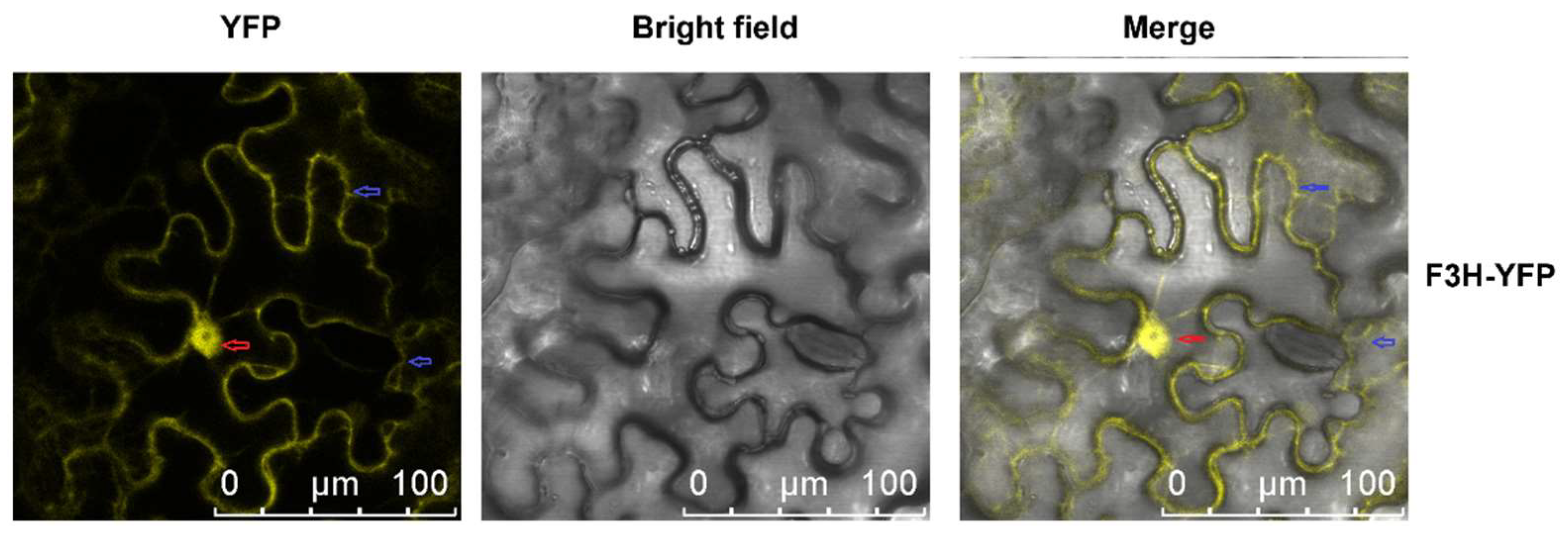
The subcellular locations of enzymes involved in secondary metabolite biosynthesis affect the utility of intermediate products and the efficiency of biosynthesis. Enzymes involved in the biosynthesis of secondary metabolites can form weakly bound, ordered complexes, which are referred to as “metabolons” [
26]. Lignins and flavonoids are products of the phenylpropanoid pathway, and both lignin and flavonoid metabolons have been reported in many plants [
26,
27]. Flavonoid metabolons appear to be organized as multi-enzyme complexes, mainly at the endoplasmic reticulum (ER) and exclusively in cytoplasm [
21]. However, flavonoids have been reported to be distributed in different cell compartments, including the cytosol, vacuole, ER, chloroplast, nucleus, and small vesicles, as well as in extracellular space, in different types of cells [
21,
22]. Despite the transport of flavonoid in cells, in situ biosynthesis of flavonoids in the nucleus has also been reported. Subcellular location analysis showed that some enzymes involved in flavonoid biosynthesis were located in both the nucleus and the cytosol. Arabidopsis AtCHS and AtCHI, CtF3H, Sm(eggplant)CHS, SmCHI, SmDFR, SmANS and
MazsF3H, in this study, were all located in both the nucleus and the cytosol [
21,
28,
29]. Meanwhile, lignin-related enzymes have been reported to be exclusively located in cytoplasm [
30]. Given that flavonoids can bind to proteins and nucleic acids and play important roles in various biological processes, including as UV protectants and modifiers of auxin transport, in situ flavonoid biosynthesis in the nucleus may serve to protect DNA from UV or to control the transcription of genes [
21,
22,
31]. It would be interesting to explore the biosynthesis mechanism of in situ flavonoid biosynthesis in the nucleus and its possible role in transcription regulation.
The regulation of structure genes involved in flavonoid biosynthesis could affect the accumulation of the products. Overexpression of
CHS and
CHI can enhance flavonoid and anthocyanin biosynthesis, and down-regulation of
CHI results in a decrease in anthocyanin content [
8,
25,
32]. The regulation of
F3H expression may have quite different results in different plants. In addition,
F3H even exhibited contrastings responses to methyl jasmonate (MeJA) in different plant phenotypes [
16]. Our results also demonstrated the complex roles of
F3H in flavonoid biosynthesis in different mulberry varieties. The expression of late anthocyanin biosynthesis genes (
UFGT,
ANS,
DFR and
CYP75B1) in fruits was much lower in non-pigmented mulberry, while the ratio of flavonols to flavonoids was higher in non-pigmented mulberry, which indicated the redirected the flux in the flavonoid pathway in non-pigmented mulberry [
33]. The relatively higher expression levels of
F3H in S1-S4 could be responsible for the accumulation of flavonols and the precursors of anthocyanins. During S5-S8, feedback regulation of
F3H may result in decreased
F3H expression levels while the accumulation of anthocyanins is still being slowly catalyzed by late anthocyanin biosynthesis enzymes, resulting in the observation of a negative correlation betwen F3H expression level and anthocyanin content. It is necessary to consider the phenotype first when trying to modify
F3H to regulate flavonoid or anthocyanin biosynthesis. In addition, the multi-level regulation of flavonoid biosynthesis in plants has also been reported, including plant hormones, transcript factors and structure genes, and protein complex. Our previous study suggested that more complex metabolons may exist in mulberry [
25]. Flavonoid biosynthesis in plants may be more complex, and more studies still need to be performed to reveal the regulation network covering different levels, especially in some featured plants.
4. Materials and Methods
4.1. Plant Materials
Samples of leaves, buds, stems, stigmata and ovaries, and fruits from
Zhongshen 1 were collected in March 2021. These samples were used for gene cloning and expression profile analysis. The samples of mulberry fruits at different development stages from
Zhongshen 1 and
LvShenZi were reported in our previous study [
25].
Morus atropurpurea variety
Zhongshen 1(
Mazs) has ripe fruits that are purple and
Morus alba var.
LvShenZi (
LSZ) has ripe fruits that are white. All of the above samples were collected from the national mulberry germplasm field of the Chinese Academy of Agricultural Sciences, Zhenjiang and immediately stored at −80 °C. At least six mulberry fruits were collected from each mulberry plant. Two-week-old seedlings of
Morus alba var.
Fengchi were used to perform virus-induced gene silencing to down-regulation of
F3H.
4.2. Isolation of RNA and cDNA Synthesis
Samples were ground with liquid nitrogen and total RNA was extracted using Plant RN38 Kit (Aidlab, Bei**g, China) according to the manual. cDNA was synthesized with the PC54-TRUEscript RT kit (Aidlab, Bei**g, China) according to the manufacturer’s protocol.
4.3. Cloning of MazsF3H
The
F3H coding region sequence was extracted from the
morus alba genome using the annotation file based on the Blast results with
AtF3H as query. The primers were designed and then synthesized using Sangon Biotech (Shanghai, China). The standard three-step PCR process was adopted with an annealing temperature of 54 °C to amplify
F3H from
Zhongshen 1. The PCR products were purified using SanPrep Column DNA Gel Extraction Kit (Sangon Biotech, Shanghai, China) and then cloned into the pMD18-T vector (Takara, Dalian, China). The sequences were deposited at GenBank (Accession number: ON055162) and named
MazsF3H. Primer information is available in
Table S2.
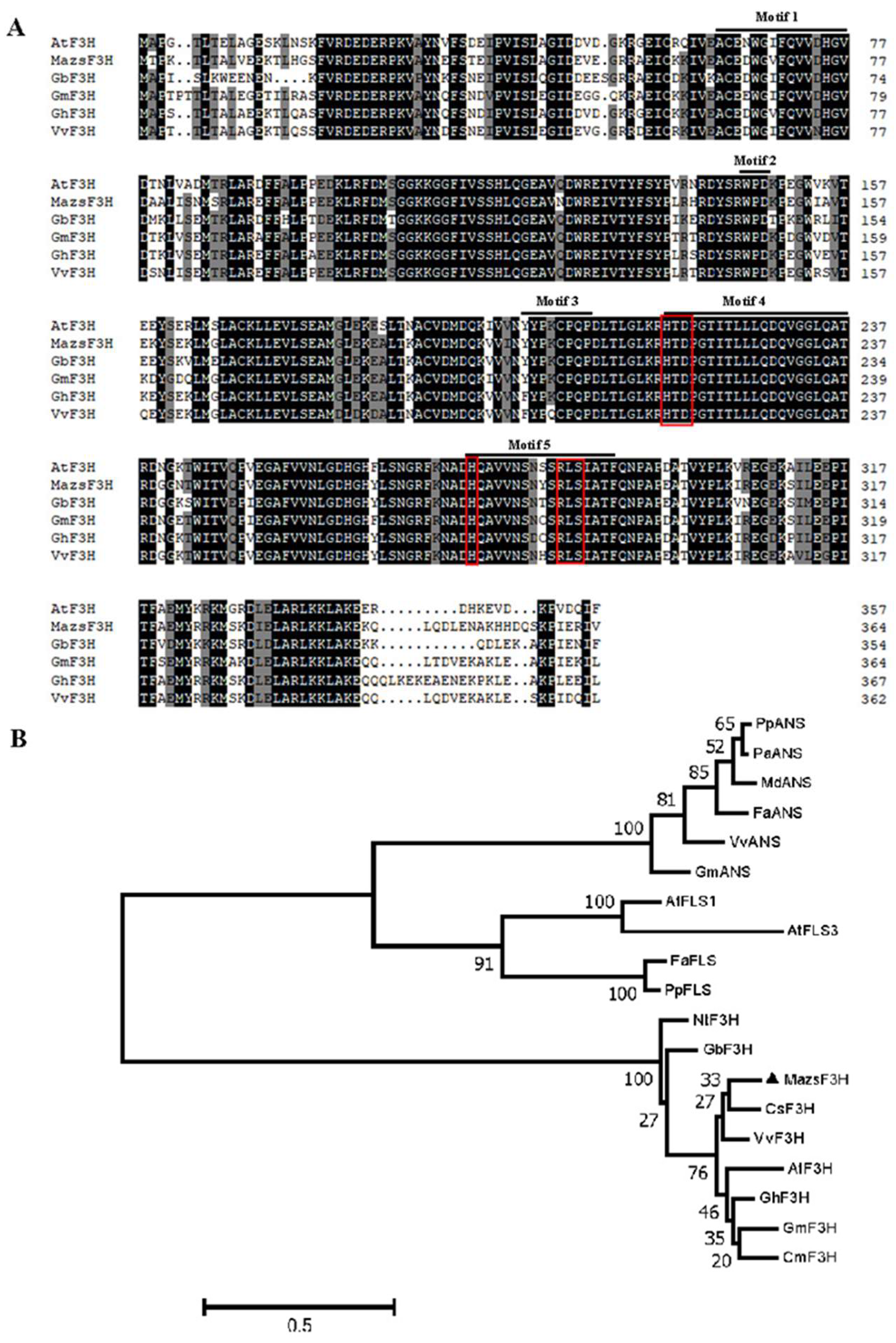
4.4. Alignment and Phylogenetic Analysis of F3Hs in Plants
MazsF3H and F3Hs from different plants, including
Arabidopsis thialana AtF3H,
Gynura bicolor GbF3H,
Garcinia mangostana GmF3H,
Gossypium hirsutum GhF3H and
Vitis vinifera VvF3H, were aligned using DNAman 8.0 (Lynnon BioSoft, QC, Canada) with the default parameters to detect the conserved motifs. In addition, alignment of 2-oxoglutarate-dependent dioxygenases (2-ODDs), including F3Hs,
FLSs and
ANSs, was performed to construct the phylogenetic tree. The phylogenetic tree was constructed using Mega 7.0 by means of the maximum-likelihood method using JTT substitution model and the G + I rates among sites model. The tree was assessed using the bootstrap** method with 1000 bootstrap replicates, and marked above nodes only if greater than 50 [
34].
4.5. Prokaryotic Expression and Purification of MazsF3H
Recombinant plasmids pET28a-MazsF3H were constructed using seamless cloning (CV1901 kit, Aidlab, Bei**g, China) and then confirmed by sequencing. Recombinant plasmids were transferred into E.coli BL21(DE3) cells and then incubated at 37 °C in LB media containing kanamycin (50 μg/mL). Finally, a concentration of 0.4 mM isopropy-β-D-thiogalactoside (IPTG) was added at OD = 0.6. Cells were collected by centrifugation at 4000 g after 4 h of incubation at 28 °C. The fusion protein was purified using Ni-NTA Sefinose Resin according to the user’s manual (BBI Life Sciences, Shanghai, China). SDS-PAGE and Bradford Protein Assay Kit (Sangon Biotech, Shanghai, China) were used to detect purified proteins and to determine the concentration of purified proteins.
4.6. Enzymatic Assay of MazsF3H
In vitro MazsF3H activity was determined as described by Si et al. (2022) with some modifications. The total 400 μL reaction mixture contained 25 mM Tris-HCl (pH 7.4), 0.4 mM naringenin, 0.5 mM dithiothreitol, 1 mM ascorbate acid, 0.2 mM ferrous sulfate, 1 mM 2-oxo-glutarate, and 10 μg or 20 μg recombinant proteins. Sample with protein that had been incubated in boiling water for 10 min were used as the negative control.
The reaction mixture was filtered using 0.22 μm PES membranes before sending them to the HPLC-VWD (Agilent 1260 infinity II, Victoria, TX, USA) system with a Poroshell EC-C18 column (4.6 mm × 150 mm, 4.0 μm, Agilent, Victoria, TX, USA). The mobile phases were methanol (A) and 1% (v/v) H3PO4 in water (B) with the following program: 0–20 min, 15% A–−60% A; 20–26 min, 60% A–0%; 26–27 min, 0% A–15% A at a flow rate of 0.8 mL/min. The substrates and products were detected at 290 nm. Standards including naringenin, dihydrokaempferol, dihydroquercetin and dihydromyricetin were purchased from Solarbio (Bei**g, China).
4.7. Expression Profile of F3H in Mulberry
qRT-PCR (quantitative real-time PCR) was performed to explore the expression of
MazsF3H in different organs, its expression pattern throughout the fruit ripening process, and in the differences in its expression in ripe fruits of different mulberry varieties using an ABI StepOnePlus™ Real-Time PCR System (Foster City, CA, USA). The primers are presented in
Table S2. Actin was used as a reference gene [
35]. GraphPad Prism 8.0 was used to visualize the qRT-PCR results and perform ANOVA.
p < 0.05 was denoted as significance. Firstly, three biological replicates were mixed, and then three technical replicates were performed, respectively, for qRT-PCR.
4.8. Subcellular Location of MazsF3H
The method used for the subcellular location of
MazsF3H was the same as that reported in our previous study [
36,
37]. The pBI121 vector with
MazsF3H gene and yellow fluorescent protein (YFP) fusion expression was constructed. The confirmed recombinant plasmids were transferred into
Agrobacterium tumefaciens strain
GV3101, which were then transferred into tobacco leaves via Agrobacterium-mediated transient transformation [
38]. The YFP fluorescence in leaves was observed using a Leica TCS SP8 confocal microscope (Leica Microsystems, Wetzlar, Germany).
4.9. Measurement of Anthocyanin and Total Flavonoid Content
The extraction and measurement of anthocyanin content were performed according to our reported methods, modified from the methods provided by Mehrtens et al. (2005) [
39]. The anthocyanin content is given in cyanidin-3-glucoside equivalents. The total flavonoid content was measured with an aluminum nitrate method, using rutin as a reference substance according to a previous study [
40].
4.10. Transient Overexpression of MazsF3H in Nicotiana Benthamiana
The recombinant plasmids
pNC-1304-35S:MazsF3H were constructed using nimble cloning [
41]. Both recombinant plasmids
pNC-1304-35S:F3H and empty vector pNC-1304-35S:GFP, as the negative control, were transformed into
Agrobacterium tumefaciens GV3101 and then transferred into
N. benthamiana leaves via Agrobacterium-mediated transient transformation, as per a previous report [
38]. Overexpression of
MazsF3H was determined using qRT-PCR by comparing the expression levels of target genes in transgenic plants with those in the negative controls.
4.11. Obtaining MazsF3H VIGS Transgenic Mulberry
Virus-induced gene silencing (VIGS) was used to obtain
MazsF3H down-regulated transgenic mulberry, in accordance with our previous report [
25,
42].
Agrobacterium tumefaciens containing recombinant plasmids
pTRV2-MazsF3H,
pTRV1 and
pTRV2 (negative control) was cultured in transient transformation buffer and then transferred into mulberry leaves by means of pressure injection. Three independent mulberry plants were injected. The knock-down efficiency for the target genes was determined by qRT-PCR 15 days after injection by comparing the transgenic plants with the negative controls and wild types. The correlation analysis and ANOVA analysis were performed using SPSS19.0.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}