1. Introduction
Tuberculosis (TB), caused by
Mycobacterium tuberculosis, is among the major infectious diseases in humans, leading to 1.5 million deaths in 2020 [
1]. The current treatment for drug-sensitive TB consists of a combination of isoniazid, rifampicin, ethambuthol, and pyrazinamide, but requires several months under strict observation. Moreover, despite efforts to end the epidemic, TB persists and antibiotic-resistant strains of
M. tuberculosis continue to emerge. In this context, it is necessary to continue efforts to develop new molecules with antimycobacterial properties and facilitators or enhancers of approved antibiotics [
2,
3].
The World Health Organization recommendations for the treatment of drug-resistant and multi-drug resistant (MDR) TB include fluoroquinolones, bedaquiline, and linezolid [
4,
5]. Bedaquiline is a new drug that was approved in 2012 for the treatment of multidrug-resistant tuberculosis with the ability to inhibit mycobacterial F1F0 ATP synthase [
6]. Linezolid is a protein synthesis inhibitor that binds to the peptidyl transferase center in the active site of 50S ribosomal subunits [
7]. First used for the treatment of infections by resistant Gram-positive bacteria, linezolid is also effective in curing patients with multiple-drug resistant (MDR) and extensive drug-resistant (XDR) TB [
8]. There are other anti-TB drugs that can be used for second-line regimens, such as fluoroquinolones, betalactams, para-aminosalicylic acid, Clofazimine, and Cycloserine. These drugs can be used in varying combinations depending on the circumstances. However, most drugs that are used for MDR TB treatments show side effects making their long-term utilization difficult. In addition,
M. tuberculosis clinical isolates resistant to bedaquiline were found after its approval, thus showing the need for other drugs in certain cases. The resurgence of MDR TB due to the COVID-19 pandemic makes it even more necessary to identify new molecules and shorten the duration of treatment [
9].
The use of drug combinations with a compound that would increase the activity of linezolid and thus reduce the amount required is among possible strategies to reduce the toxicity of linezolid. Combination therapeutics are also excellent strategies to limit the emergence of antibiotic resistance and thus extend the life of antibiotics [
10]. We have previously developed a whole-cell screening method coupled with a resazurin reduction as a marker of cellular viability to identify molecules with anti-mycobacterial activity [
11]. In order to identify the potential target or the mode of action of the hit compounds, it is possible to use the genome-wide transcriptional response of
Mycobacterium exposed to the compound [
12] or the whole-genome sequencing of resistant mutants [
13].
In this study, we undertook whole-cell screening in the presence of subinhibitory concentrations of linezolid coupled with whole-genome sequencing of spontaneous resistant mutants to identify synergistic or compatible combination treatments and their target. Among 52,000 chemical compounds of various families, 15 compounds similar to or different from known antibiotics were found with a minimal inhibitory concentration <2 µg/mL. Here, we describe one small molecule with a novel structure that targets the essential lipid transmembrane transporter MmpL3, required for the export of the lipid trehalose monomycolate (TMM) [
14,
15]. Then, the toxicity of this compound to mammalian cells was determined, as well as its efficacy in the therapy of experimentally
M. bovis BCG infected
Galleria larvae.
3. Materials and Methods
3.1. Drugs and Reagent Preparation
The commercial drugs isoniazid, rifampicin, and linezolid were purchased from Sigma, bedaquiline and nigericin from TargetMol Chemicals Inc, Boston, USA, and valinomycin from Shanghai Acmec Biochemical Co., Ltd, Shanghai, China. Antibiotic solutions were prepared at a concentration of 5 mg/mL in dimethyl sulfoxide (DMSO) from Sangon Biotech Co., Ltd, Shanghai, China.
The 52,000 compounds from Topscience Co., Shanghai, China, to be screened were dissolved in 100% DMSO. Lead compound 1 showed 98.6% purity in HPLC analysis. Resazurin sodium salt powder (BBI Life Sciences, Shanghai, China) was prepared at 0.01% (w/v) in distilled water, filter sterilized, and stored at 4 °C for up to two weeks.
3.2. Strains and Growth Condition
M. bovis BCG (strain 1173P2), M. marinum (ATCC BAA-535), M. smegmatis mc2155, M. abscessus (ATCC 19977), M. aurum, M. avium (ATCC 25291), and M. tuberculosis H37Rv (ATCC 27294) were obtained from the collection of the Institut Pasteur. M. tuberculosis clinical isolates, including three susceptible, three Rif-resistant and one MDR strain were collected from hospitals in China. Mycobacteria were cultured in Middlebrook 7H9 broth containing 10% (v/v) ADC enrichment (albumin, dextrose, catalase; Becton Dickinson, Franklin Lakes, USA), 0.05% glycerol (Sangon Biotech, Shanghai, China), and 0.05% Tween 80 (Sangon Biotech, Shanghai, China), or Middlebrook 7H11 agar medium supplemented with 10% (v/v) OADC enrichment (oleic acid, albumin, dextrose, catalase; Becton Dickinson, Franklin Lakes, USA). All mycobacteria were cultured under aerobic conditions at 37 °C, except for M. marinum, which was cultured at 30 °C. E. coli ATCC 25922 and S. aureus ATCC 25923 were cultivated at 37 °C in LB medium.
3.3. Screening and MIC and MBC Determination
Screening of a chemical library composed of 52,000 compounds from Top Science was performed at the People’s Hospital of Shenzhen. The small chemical compounds were screened at 2 μg/mL (which corresponds to an average concentration of 5 to 6 μM) in the presence of 0.15 µg/mL linezolid. Each compound was prepared in 50 µL Middlebrook 7H9 broth in a 96-well plate and a 50 µL aliquot of M. bovis BCG suspension then added to each well. The plates were incubated at 37 °C for two weeks to allow growth of the bacteria. The antimycobacterial activity of compounds was determined using the resazurin-reduction assay. A change in color from blue to pink indicated bacterial growth. DMSO was used as a positive control to define growth of the bacteria without any compound (100% viability) and rifampicin was used as a negative control to kill all bacteria (0% viability). Primary hits were filtered using an activity cut-off (MIC < 4 µg/mL against M. bovis BCG) and by selecting for drug-like properties.
Determinations of MICs were performed by the microdilution method in 96-well plates, with each well containing 100 µL bacterial suspension with a serial dilution of the compound. The MICs were defined as the lowest concentration of compound that inhibited bacterial growth and thus prevented the change in color of resazurin. Pink wells indicated bacterial growth and blue wells no bacterial growth. Isoniazid and rifampicin were used as standards.
MBCs were determined by transferring 100 µL from each of the wells from the MIC assays (starting from the MIC for each compound) onto 7H11 plates. Plates were incubated at 37 °C for two days (M. smegmatis) or three weeks (M. bovis). The MBC was defined as the first plate (lowest concentration) yielding no growth.
3.4. Dose–Response Curves
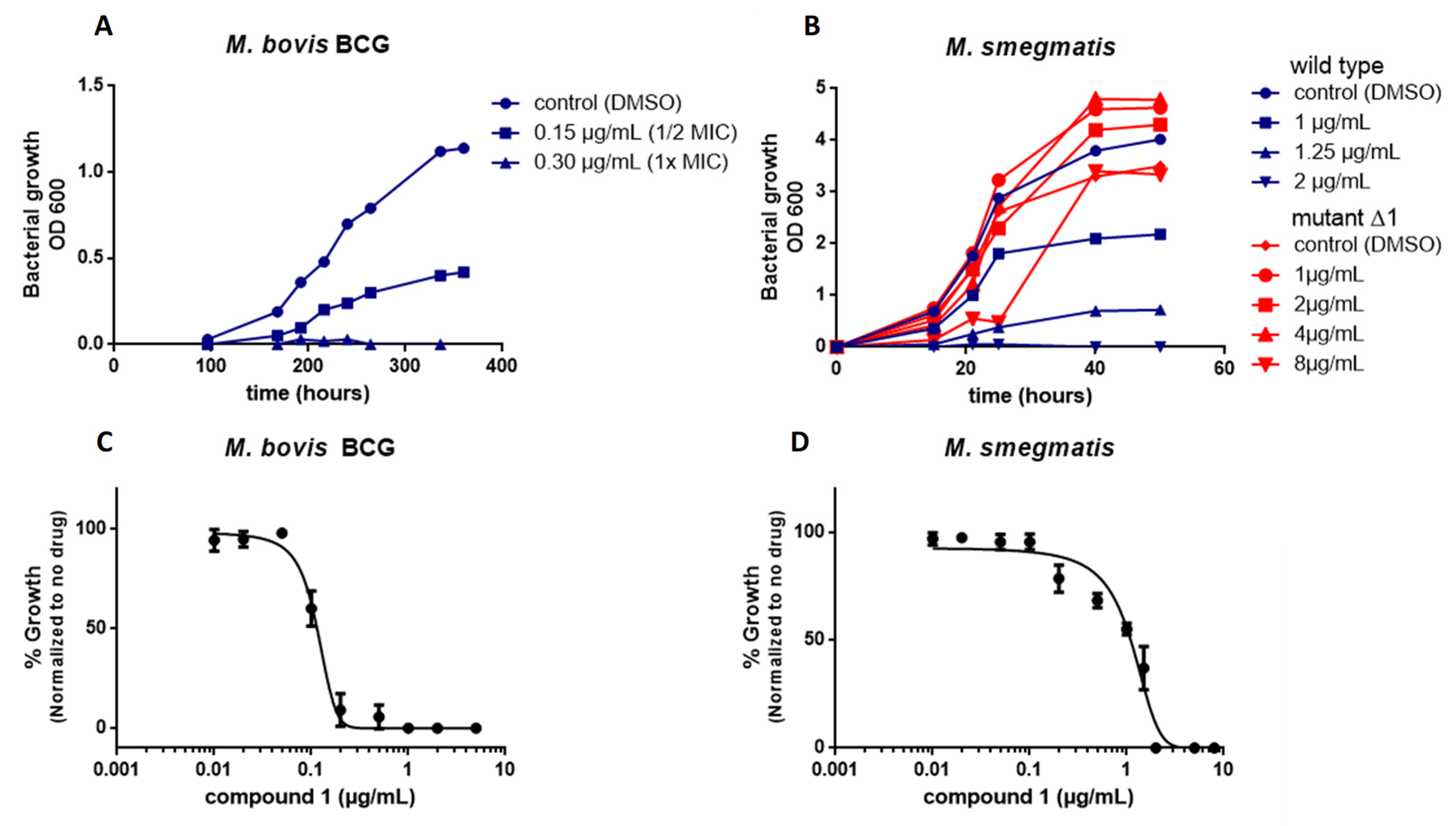
M. smegmatis and M. bovis BCG were grown in 7H9 medium to an optical density at 600 nm of 0.5 to 1.0 and the cultures then diluted to an OD600 of 0.05 in 7H9 medium. Compounds were tested at concentrations between 0.01 and 5 µg/mL. The controls included DMSO and 2 µg/mL rifampicin. The antimicrobial potency of the compounds was evaluated at 600 nm by comparison with DSMO-treated (drug free, 100% growth) and rifampicin-treated bacterial suspensions (0%). Dose–response assays were conducted in triplicate and the curves created using GraphPad Prism 6 software (San Diego, CA, USA, version 6.0.1).
3.5. Kill Kinetics
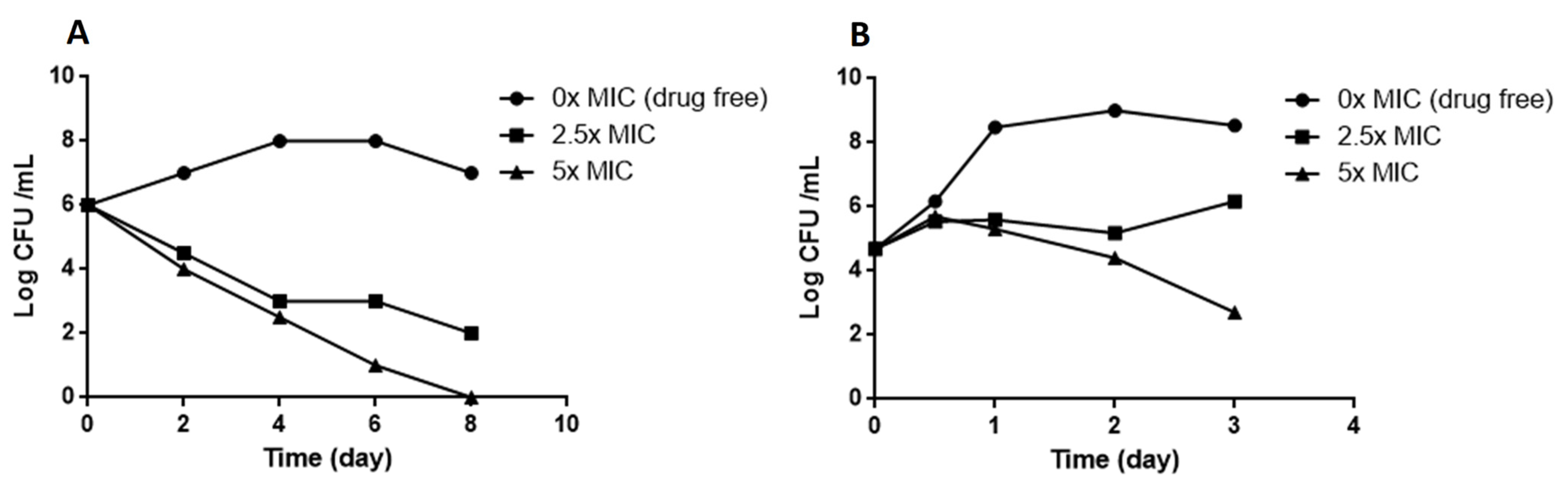
M. smegmatis and M. bovis BCG were cultured in 7H9 broth to log phase (105–106 CFU/mL), various concentrations of compound 1 in 7H9 (final concentration 0.5% DMSO) added, and the cultures incubated at 37 °C. The untreated control was 0.5% DMSO. The number of viable bacteria was determined by serial dilution and plating on Middlebrook 7H11 plus 10% vol/vol OADC. CFUs were counted after three days for M. smegmatis and 3 to 4 weeks for M. bovis BCG.
3.6. Generation and Analysis of Resistant Mutants
Resistant mutants were isolated by plating 109 cells of M. smegmatis or M. bovis BCG (in 300 µL) onto plates containing 5 mL 7H11 medium with the 4×, 8×, and 16× liquid MIC of the compound. Resistant colonies were inoculated in 7H9 liquid medium containing the 2× MIC of the compound. The MICs of selected colonies were then determined by serial dilution. The MIC determinations were performed in triplicate for each strain of bacteria. The rate of resistance mutations was calculated as the ratio of CFU in the absence of the compound and CFU in presence of the compound.
3.7. Genome Sequencing and the Identification of Polymorphisms
The genomic DNA of selected mutants was extracted and purified using EZ-10 Spin columns from bacterial genomic DNA Mini-Prep Kits (Bio Basic, Amherst, MA, USA), sequenced, and analyzed for SNPs. Genome sequencing was performed by Sangon Biotech Co., Ltd, Shanghai, China. The genomes were sequenced using an Illumina HiSeq 2500 platform (Illumina, San Diego, CA, USA) and then quality-filtered and assembled using SPAdes Genome Assembler (St. Petersburg, Russia, version 3.12.0). The resulting reads were mapped to the
M. smegmatis MC2 155 reference genome and mutations were identified using the Snippy pipeline (
https://github.com/tseemann/snippy) (accessed on 16 July 2021).
The sequences assembled in the present study can be accessed through GenBank (NCBI) using the accession codes PRJNA766507.
3.8. Checkerboard Synergy Assay
Drug interactions between the identified compound and other antimycobacterial drugs were assessed with M. bovis BCG and M. smegmatis using the checkerboard microdilution method and resazurin assay as a viability marker.
Briefly, 50 µL 7H9 medium was distributed into each well of 96-well microdilution plates. Serial two-fold dilutions of each drug to at least double the MIC were prepared prior to testing. Compound 1 was serially diluted along the ordinate and the second drug along the abscissa. Each well was then inoculated with 50 µL of a bacterial inoculum of 105 CFU/mL and the plates were incubated at 37 °C for seven days under aerobic conditions. The experiment was carried out in triplicate.
The fractional inhibitory concentration index (FICI) was determined in a 96-well plate and calculated by adding the FIC
A (MIC of compound A in the presence of compound B)/(MIC of compound A alone) and the FIC
B (MIC of compound B in the presence of compound A)/(MIC of compound B alone). Synergy was defined as an FICI value ≤ 0.5, while the values 0.5 < FICI ≤ 1 correspond to additivity; 1 < FICI ≤ 4 is indifferent; FICI > 4 indicate antagonism [
56,
57].
3.9. Docking Studies
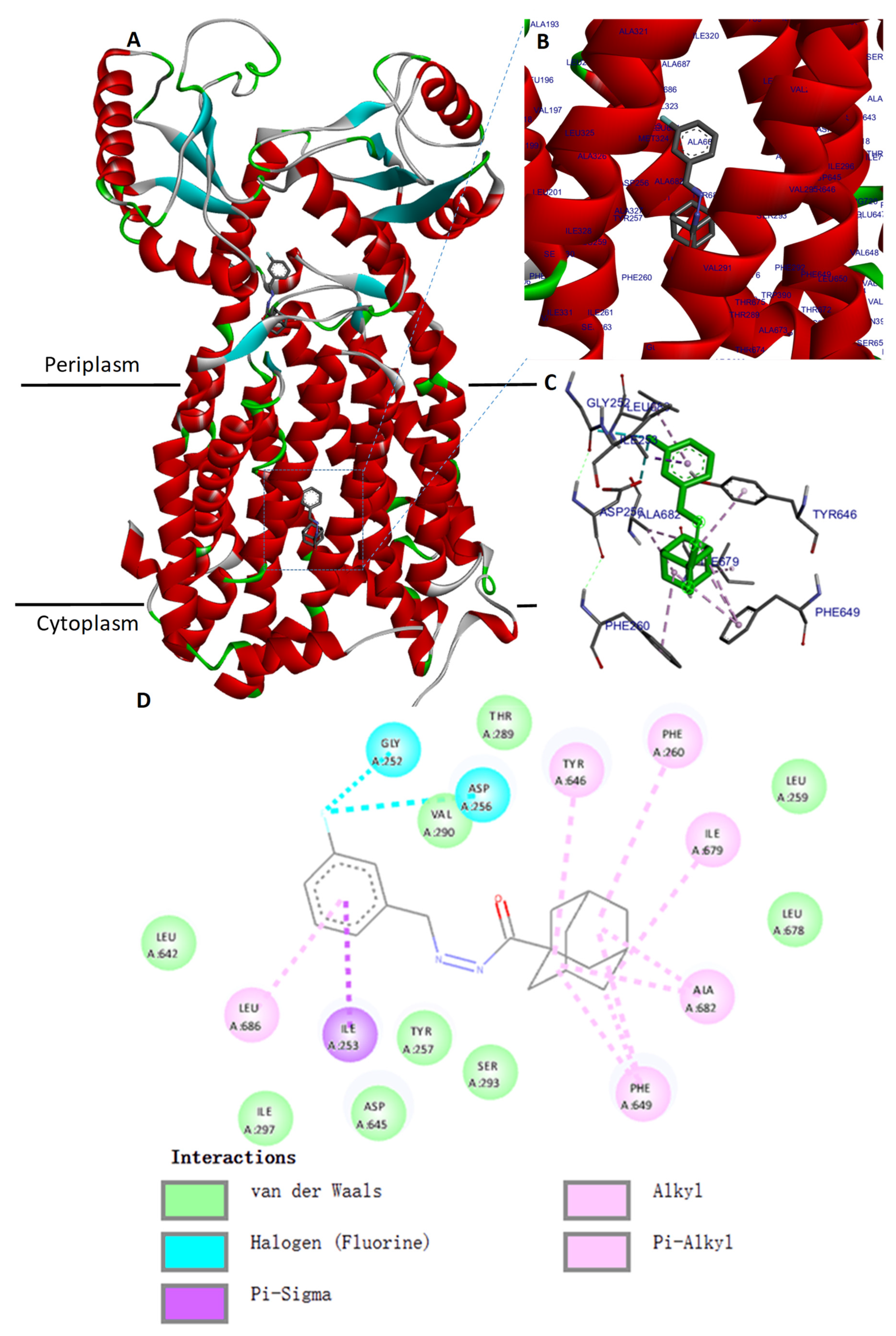
Docking of compound 1 was carried out using the crystal structure of MmpL3 from
M. smegmatis (PDB ID: 6AJH) obtained from the Protein Data Bank. This protein was considered as the target, and was prepared by adding hydrogens, calculated Gasteiger charges, and the complexed ligands were manually removed using the AutodockVina program. Compound
1, considered as the ligand, was sketched using Marvin Sketch tools from ChemAxon (Budapest, Hongary) and converted into 3D using Discovery Studio Analyzer. The docking simulation was performed using Autodock Vina. A grid box was created with 50 × 50 × 50 points, with a resolution of 0.375 Å. The coordinates of the grid center were x 37.00, y 4.00, and z −22.00. The Lamarckian genetic algorithm was used to run 100 dockings. The docked conformations were ranked according to the docking free energy. Information was collected and ligand binding site atoms were visualized using Biovia Discovery Studio Analyzer. The same procedure was performed using AU1235 as the control ligand to validate the docking protocol. Then, compound
1 and AU1235 were docked in the same way against MmpL3 from
M. tuberculosis (PDB ID: 7NVH) [
47].
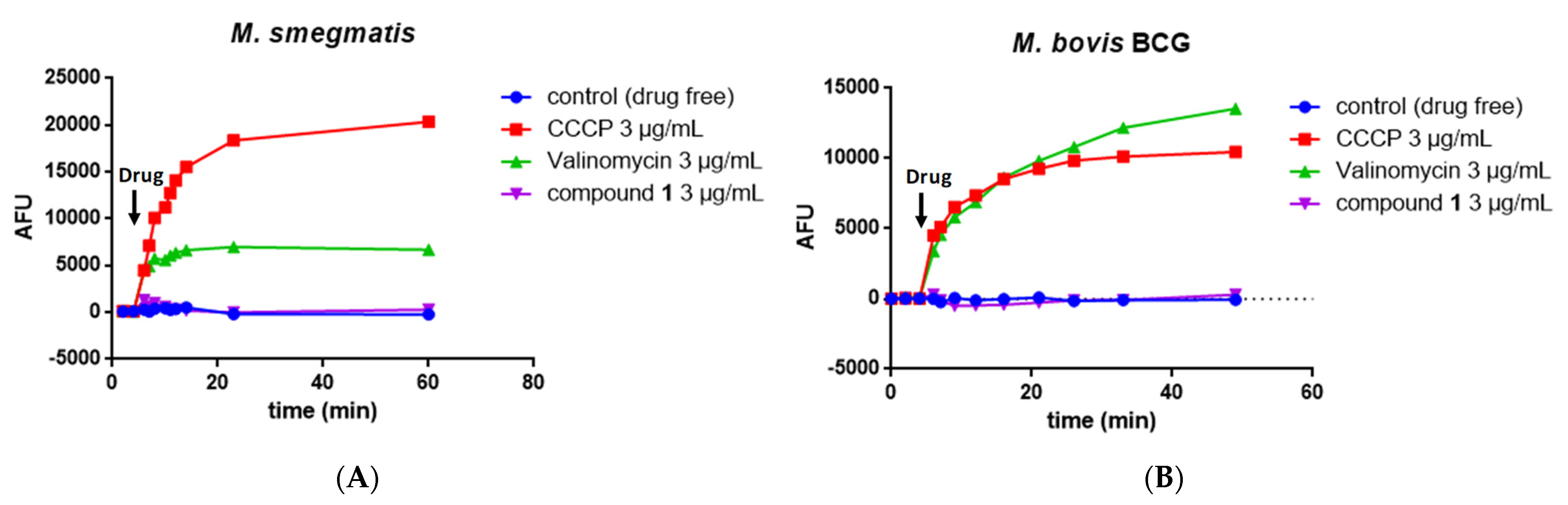
3.10. Measurement of the Mycobacterial Transmembrane Potential ΔΨ
Exponentially growing M. bovis BCG or M. smegmatis were harvested by centrifugation, washed once with buffer containing 5 mM Hepes, 5 mM glucose, and 0.05% Tween (pH 7.2), resuspended in the same buffer containing 2 μM DiSC3(5), and incubated for 2 h at 37 °C. Then, suspensions were dispensed into costar white flat-bottom 96-well plates (100 μl/well) and treated with compounds, CCCP, or valinomycin. Fluorescence was monitored (excitation wavelength, 622 nm; emission wavelength, 670 nm) on a Tecan Spark 10 M microplate reader at room temperature. Wells containing cells in the presence of DiSC3(5) and DMSO served as controls. All assays were performed at least twice (a representative assay is shown).
3.11. Measurement of the Mycobacterial Chemical Proton Potential ΔpH
M. smegmatis or M. bovis BCG were labeled with 20 µM of the fluorophore BCECF-AM (Invitrogen) in 0.1 M Hepes buffer, pH 7.0, for 30 min at 37 °C. Cells were pelleted by centrifugation and re-suspended in 0.1 M Hepes at various pHs (6.0–8.0) in the presence of 20 μM nigericin to generate a standard curve. Fluorescence emission (λem 525 nm) intensities of intracellular BCECF were measured following excitation at λex 488 and 440 nm in a Tecan Spark 10 M microplate reader. The effect on intracellular pH was studied after the treatment of Mycobacterium cells with various concentrations of compound 1 in 0.1 M Hepes at pH 6.8. DMSO was used as a negative control and CCCP and nigericin as positive controls.
3.12. Adme Prediction
An ADME (adsorption, distribution, metabolism, and excretion) prediction was performed in silico using the web tool SWISS-ADME (
https://www.swissadme.ch) (accessed on 17 October 2022).
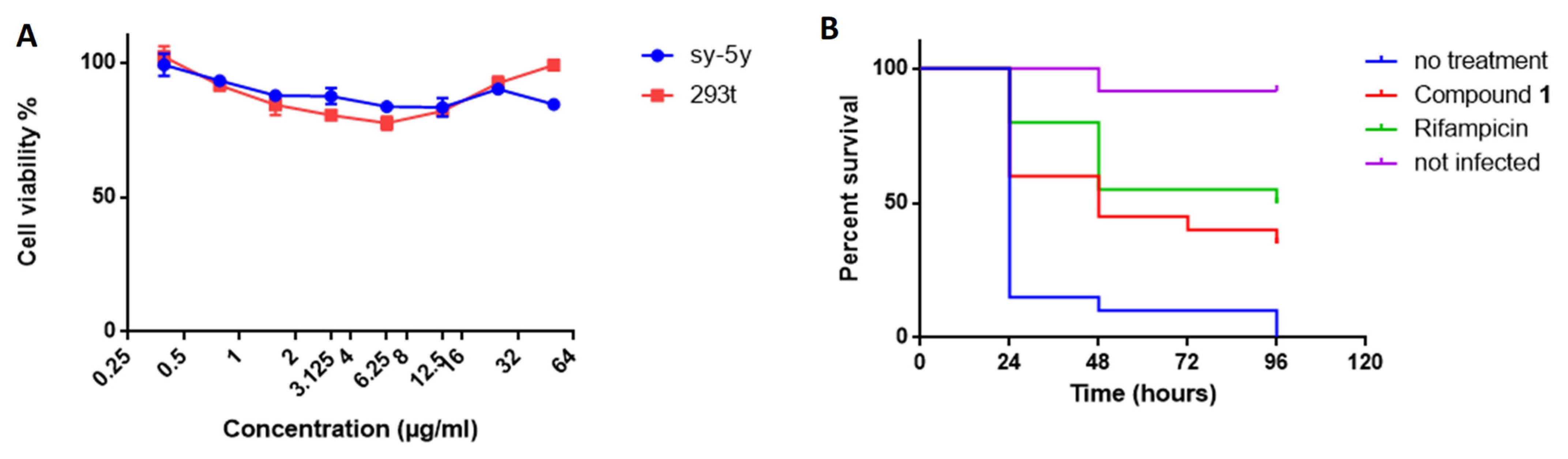
3.13. Cytotoxicity
The cytotoxicity of compound 1 was assessed on two different mammalian cell lines (Sh-SY5Y and HEK 293T) by the MTT colorimetric assay using tetrazolium reduction. Cell suspensions were cultivated in RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS) and incubated at 37 °C in a 5% CO2 atmosphere. The cytotoxicity assays were performed in 96-well microplates, adding 100 µL of culture with 4 × 104 cells/mL to each well. On the next day, cells were treated with the compounds prepared in DMSO and serially diluted from 50 to 0.1 µg/mL. Each drug was tested in triplicate. The plates were examined after 48 h of incubation, according to the manufacturer’s instructions. The percentage viability was calculated by comparison with the control without treatment.
3.14. Infection of Galleria Mellonella and Drug Treatment
Larvae were purchased from Bei**g KuoyeTianyuan Biotechnology C., Ltd, Bei**g, China. Healthy, high-mobility, ~300-mg cream-colored larvae were used for the infection experiments. Injections into the hemocoel via the last left proleg were performed with a 10-μL Hamilton syringe. For infection, 10 µL of a
M. bovis BCG (10
7 CFU) suspension in PBS, 0.05% Tween was injected [
53]. For treatment, 10 µL of compound (rifampicin or compound 1) diluted in 6% DMSO at a dose of 10 mg/kg was injected 2 h after infection (
n = 20). The control group of infected larvae was injected with 10 µL 6% DMSO (
n = 20) and the control group of uninfected larvae (
n = 20) with 10 µL PBS, 0.05% Tween. Larvae were then incubated at 37 °C in Petri dishes (5 per box) in the dark without food.
Survival of the infected larvae (n = 20 per group) following treatment was monitored every 24 h for 96 h. Larvae were considered dead when they were unresponsive to touch. Kaplan–Meier survival curves were used to compare the distribution of survival of each group of larvae.
4. Conclusions
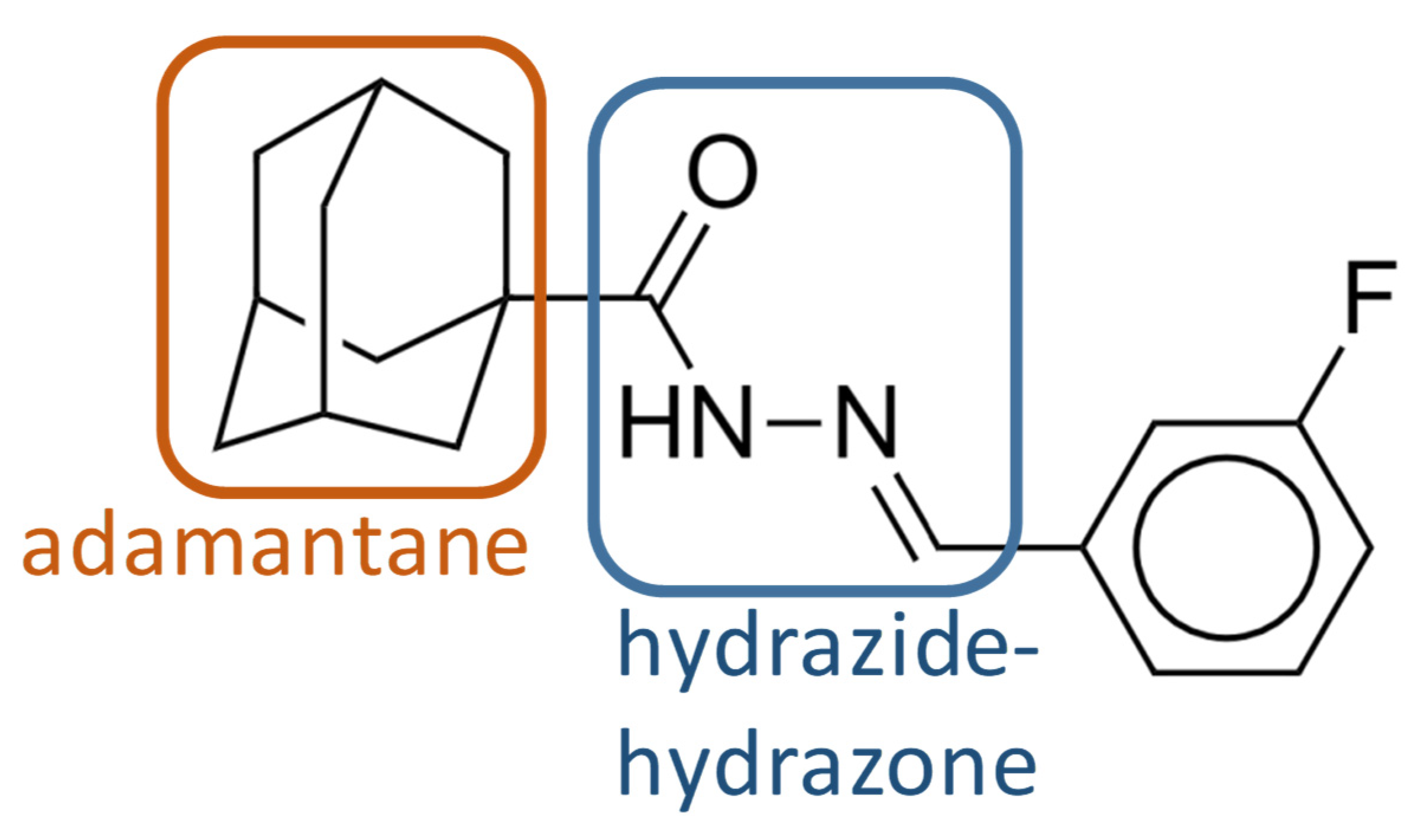
The emergence of drug-resistant TB is a major global health challenge. Controlling the disease will require new molecules that can be used in combination with approved drugs to improve and shorten treatment time. Here, phenotypic screening of a small-molecule chemical library allowed the identification of several molecules with anti-mycobacterial activity. Among them, we identified compound 1 as a probable inhibitor of MmpL3, with a new scaffold containing an adamantane moiety and a hydrazide–hydrazone moiety. Compound 1 showed antimycobacterial activity against M. smegmatis, M. bovis BCG, and M. tuberculosis H37Rv. It was also active against a panel of clinical drug-sensitive, Rif-R, and MDR M. tuberculosis strains, with MIC values of 0.2 to 0.4 µg/mL and moderately against the nontuberculous mycobacterial species M. abscessus (MIC value of 12.5 µg/mL).
Whole-genome sequencing of spontaneous mutants resistant to compound
1 allowed us to identify MmpL3 as the putative target of compound
1. MmpL3 is an inner membrane protein that transports mycolic acids in the form of trehalose monomycolates (TMMs) across the membrane. We generated independent compound
1-resistant mutants with a F649L mutation localized in the transmembrane segment of MmpL3, inside the proton-translocation channel. Amino acid changes at F649 in
M. smegmatis correspond to F644 in
M. tuberculosis, which have been reported previously for other MmpL3 inhibitors [
13,
31,
44,
58]. Molecular docking suggested that compound
1 could also binds within the binding pocket of
M. smegmatis MmpL3 and interacts with the two Asp-Tyr pairs (Asp256-Tyr646 and Asp645-Tyr257) involved in proton translocation which allow the energy necessary for the substrate translocation [
44,
45], although the docking of compound
1 against the Cryo-EM structure of
M. tuberculosis MmpL3 did not reveal an interaction within the binding pocket in the same way as in
M. smegmatis, but only in the cavity at the center of the periplasmic domain.
Moreover, the mode of action of MmpL3 inhibitors is debated because the activity of MmpL3 is also linked to the proton motive force (PMF) of the inner membrane. Several studies have proposed that certain inhibitors indirectly target MmpL3 and disrupt the membrane potential [
51,
58], whereas others do not [
29]. If compound
1 targeted MmpL3, we also found that this compound did not affect the inner membrane PMF.
We show that compound 1 acts synergistically in combination with linezolid against M. bovis BCG and M. smegmatis but not against M. tuberculosis. An enhancer that increases the activity of linezolid could allow a reduction in the dose of linezolid during treatment and thus reduce adverse effects. Future works with other compounds showing similarity with compound 1 may provide useful information.
We investigated the effect of compound 1 on a G. mellonella larvae model of infection to evaluate its activity in vivo. Compound 1 showed no toxicity towards the larvae and a protective effect against M. bovis BCG. This is the first time that an inhibitor targeting MmpL3 has been used in G. mellonella larvae, validating the suitability of G. mellonella larvae as an inexpensive and efficient in vivo model for the characterization of new antimycobacterial compounds. Based on our results, we consider compound 1 to have promising therapeutic potential.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}