

Molecular Mechanisms and Applications of N-Acyl Homoserine Lactone-Mediated Quorum Sensing in Bacteria
 , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. AHL Signal
2.1. AHL Diversity
2.2. AHL Production
2.3. AHL Synthesis Protein Biomolecules
2.3.1. LasI
2.3.2. RhlI
2.3.3. CviI
2.3.4. TraI
3. AHL-Dependent Transcription Regulator Biomolecules
3.1. LasR
3.2. RhlR
3.3. QscR
3.4. TraR
3.5. AHL Production
3.6. SdiA
4. Applications of AHL-Dependent Quorum Sensing
4.1. Human Health
4.2. Controlling Plant Diseases
4.3. Plant Growth Promotion and Defense
4.4. Biofouling
4.5. Bioremediation
4.6. Biosensor Development
4.7. Cancer
5. Conclusion and the Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boo, A.; Amaro, R.L.; Stan, G.-B. Quorum sensing in synthetic biology: A review. Curr. Opin. Syst. Biol. 2021, 28, 100378. [Google Scholar] [CrossRef]
- Azimi, S.; Klementiev, A.D.; Whiteley, M.; Diggle, S.P. Bacterial quorum sensing during infection. Annu. Rev. Microbiol. 2020, 74, 201–219. [Google Scholar] [CrossRef]
- Acet, Ö.; Erdönmez, D.; Acet, B.Ö.; Odabaşı, M. N-Acyl Homoserine lactone molecules assisted quorum sensing: Effects consequences and monitoring of bacteria talking in real life. Arch. Microbiol. 2021, 203, 3739–3749. [Google Scholar] [CrossRef] [PubMed]
- Bottomley, M.J.; Muraglia, E.; Bazzo, R.; Carfì, A. Molecular insights into quorum sensing in the human pathogen Pseudomonas aeruginosa from the structure of the virulence regulator LasR bound to its autoinducer. J. Biol. Chem. 2007, 282, 13592–13600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chbib, C. Impact of the structure-activity relationship of AHL analogues on quorum sensing in Gram-negative bacteria. Bioorg. Med. Chem. 2020, 28, 115282. [Google Scholar] [CrossRef] [PubMed]
- Ruhal, R.; Kataria, R. Biofilm patterns in Gram-positive and Gram-negative bacteria. Microbiol. Res. 2021, 251, 126829. [Google Scholar] [CrossRef]
- Egland, K.A.; Greenberg, E.P. Quorum sensing in Vibrio Fischeri: Analysis of the LuxR DNA binding region by alanine-scanning mutagenesis. J. Bacteriol. 2001, 183, 382–386. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.; Lu, R.; Liu, F.; Ye, J.; Zhao, J.; Wang, F.; Yang, M. Identification of LuxR family regulators that integrate into quorum sensing circuit in Vibrio parahaemolyticus. Front. Microbiol. 2021, 12, 691842. [Google Scholar] [CrossRef]
- Sırıken, B.; Öz, V.; Erol, İ. Quorum sensing systems, related virulence factors, and biofilm formation in Pseudomonas aeruginosa Isolated from Fish. Arch. Microbiol. 2021, 203, 1519–1528. [Google Scholar] [CrossRef]
- Naga, N.G.; El-Badan, D.E.; Rateb, H.S.; Ghanem, K.M.; Shaaban, M.I. Quorum sensing inhibiting activity of cefoperazone and its metallic derivatives on Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2021, 11, 716789. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Bassler, B.L. Bacterial quorum sensing in complex and dynamically changing environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Q.; Dong, D.; Hu, H.; Wu, B.; Ren, H. AHLs-mediated quorum sensing threshold and its response towards initial adhesion of wastewater biofilms. Water Res. 2021, 194, 116925. [Google Scholar] [CrossRef] [PubMed]
- Prescott, R.D.; Decho, A.W. Flexibility and adaptability of quorum sensing in nature. Trends Microbiol. 2020, 28, 436–444. [Google Scholar] [CrossRef]
- Baltenneck, J.; Reverchon, S.; Hommais, F. Quorum sensing regulation in phytopathogenic bacteria. Microorganisms 2021, 9, 239. [Google Scholar] [CrossRef]
- Von Bodman, S.B.; Willey, J.M.; Diggle, S.P. Cell-cell communication in bacteria: United we stand. J. Bacteriol. 2008, 190, 4377–4391. [Google Scholar] [CrossRef] [Green Version]
- Ampomah-Wireko, M.; Luo, C.; Cao, Y.; Wang, H.; Nininahazwe, L.; Wu, C. Chemical probe of AHL modulators on quorum sensing in Gram-negative bacteria and as antiproliferative agents: A review. Eur. J. Med. Chem. 2021, 226, 113864. [Google Scholar] [CrossRef]
- Churchill, M.E.A.; Chen, L. Structural basis of acyl-homoserine lactone-dependent signaling. Chem. Rev. 2011, 111, 68–85. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Li, W.; Liu, K.; Li, H.; Lan, X. C4-HSL Aptamers for blocking qurom sensing and inhibiting biofilm formation in Pseudomonas aeruginosa and Its structure prediction and analysis. PLoS ONE 2019, 14, e0212041. [Google Scholar] [CrossRef]
- Marketon, M.M.; Gronquist, M.R.; Eberhard, A.; González, J.E. Characterization of the sinorhizobium meliloti SinR/SinI locus and the production of novel N-Acyl homoserine lactones. J. Bacteriol. 2002, 184, 5686–5695. [Google Scholar] [CrossRef]
- Santajit, S.; Seesuay, W.; Mahasongkram, K.; Sookrung, N.; Pumirat, P.; Ampawong, S.; Reamtong, O.; Chongsa-Nguan, M.; Chaicumpa, W.; Indrawattana, N. Human single-chain variable fragments neutralize Pseudomonas aeruginosa quorum sensing molecule, 3O-C12-HSL, and prevent cells from the HSL-mediated apoptosis. Front. Microbiol. 2020, 11, 1172. [Google Scholar] [CrossRef] [PubMed]
- Thiel, V.; Kunze, B.; Verma, P.; Wagner-Döbler, I.; Schulz, S. New structural variants of homoserine lactones in bacteria. Chembiochem 2009, 10, 1861–1868. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Allen, M.S.; Simpson, M.L.; Sayler, G.S.; Cox, C.D. Direct quantification of N-(3-oxo-hexanoyl)-L-homoserine lactone in culture supernatant using a whole-cell bioreporter. J. Microbiol. Methods 2007, 68, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Beaber, J.W.; Moré, M.I.; Fuqua, C.; Eberhard, A.; Winans, S.C. Analogs of the autoinducer 3-oxooctanoyl-homoserine lactone strongly inhibit activity of the TraR protein of Agrobacterium tumefaciens. J. Bacteriol. 1998, 180, 5398–5405. [Google Scholar] [CrossRef] [Green Version]
- Morohoshi, T.; Ogasawara, Y.; **e, X.; Hamamoto, H.; Someya, N. Genetic and biochemical diversity for N-acylhomoserine lactone biosynthesis in the plant pathogen Pectobacterium carotovorum subsp. Carotovorum. Microbes Environ. 2019, 34, 429–435. [Google Scholar] [CrossRef] [Green Version]
- Gambello, M.J.; Iglewski, B.H. Cloning and characterization of the Pseudomonas aeruginosa LasR gene, a transcriptional activator of elastase expression. J. Bacteriol. 1991, 173, 3000–3009. [Google Scholar] [CrossRef] [Green Version]
- Rather, M.A.; Saha, D.; Bhuyan, S.; Jha, A.N.; Mandal, M. Quorum quenching: A drug discovery approach against Pseudomonas aeruginosa. Microbiol. Res. 2022, 264, 127173. [Google Scholar] [CrossRef]
- Dunphy, G.; Miyamoto, C.; Meighen, E. A Homoserine lactone autoinducer regulates virulence of an insect-pathogenic bacterium, Xenorhabdus nematophilus (Enterobacteriaceae). J. Bacteriol. 1997, 179, 5288–5291. [Google Scholar] [CrossRef] [Green Version]
- Roche, D.M.; Byers, J.T.; Smith, D.S.; Glansdorp, F.G.; Spring, D.R.; Welch, M. Communications blackout? Do N-acylhomoserine-lactone-degrading enzymes have any role in quorum sensing? Microbiology 2004, 150, 2023–2028. [Google Scholar] [CrossRef] [Green Version]
- Cruz, R.L.; Asfahl, K.L.; Van den Bossche, S.; Coenye, T.; Crabbé, A.; Dandekar, A.A. RhlR-regulated acyl-homoserine lactone quorum sensing in a cystic fibrosis isolate of Pseudomonas aeruginosa. MBio 2020, 11, e00532-20. [Google Scholar] [CrossRef]
- Morohoshi, T.; Kato, M.; Fukamachi, K.; Kato, N.; Ikeda, T. N-acylhomoserine lactone regulates violacein production in Chromobacterium violaceum type strain ATCC 12472. FEMS Microbiol. Lett. 2008, 279, 124–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddela, N.R.; Sheng, B.; Yuan, S.; Zhou, Z.; Villamar-Torres, R.; Meng, F. Roles of quorum sensing in biological wastewater treatment: A critical review. Chemosphere 2019, 221, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Suppiger, A.; Schmid, N.; Aguilar, C.; Pessi, G.; Eberl, L. Two quorum sensing systems control biofilm formation and virulence in members of the burkholderia cepacia complex. Virulence 2013, 4, 400–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesh, P.S.; Vishnupriya, S.; Vadivelu, J.; Mariappan, V.; Vellasamy, K.M.; Shankar, E.M. Intracellular survival and innate immune evasion of Burkholderia cepacia: Improved understanding of quorum sensing-controlled virulence factors, biofilm, and inhibitors. Microbiol. Immunol. 2020, 64, 87–98. [Google Scholar] [CrossRef]
- Ulrich, R.L.; Deshazer, D.; Hines, H.B.; Jeddeloh, J.A. Quorum sensing: A transcriptional regulatory system involved in the pathogenicity of Burkholderia mallei. Infect. Immun. 2004, 72, 6589–6596. [Google Scholar] [CrossRef] [Green Version]
- Cataldi, T.R.I.; Bianco, G.; Abate, S.; Losito, I. Identification of unsaturated N-acylhomoserine lactones in bacterial isolates of Rhodobacter sphaeroides by liquid chromatography coupled to electrospray ionization-hybrid linear ion trap-fourier transform ion cyclotron resonance mass spectrometry. Rapid Commun. Mass Spectrom. 2011, 25, 1817–1826. [Google Scholar] [CrossRef]
- Velázquez-Sánchez, C.; Espín, G.; Peña, C.; Segura, D. The modification of regulatory circuits involved in the control of polyhydroxyalkanoates metabolism to improve their production. Front. Bioeng. Biotechnol. 2020, 8, 386. [Google Scholar] [CrossRef]
- Montebello, A.N.; Brecht, R.M.; Turner, R.D.; Ghali, M.; Pu, X.; Nagarajan, R. Acyl-ACP substrate recognition in Burkholderia mallei BmaI1 acyl-homoserine lactone synthase. Biochemistry 2014, 53, 6231–6242. [Google Scholar] [CrossRef] [Green Version]
- Billot, R.; Plener, L.; Jacquet, P.; Elias, M.; Chabrière, E.; Daudé, D. Engineering acyl-homoserine lactone-interfering enzymes toward bacterial control. J. Biol. Chem. 2020, 295, 12993–13007. [Google Scholar] [CrossRef]
- Lutter, E.; Lewenza, S.; Dennis, J.J.; Visser, M.B.; Sokol, P.A. Distribution of quorum-sensing genes in the Burkholderia cepacia complex. Infect. Immun. 2001, 69, 4661–4666. [Google Scholar] [CrossRef]
- Zaitseva, Y.V.; Koksharova, O.A.; Lipasova, V.A.; Plyuta, V.A.; Demidyuk, I.V.; Chernin, L.S.; Khmel, I.A. SprI/SprR quorum sensing system of Serratia proteamaculans 94. Biomed Res. Int. 2019, 2019, 3865780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellington Miranda, S.; Cong, Q.; Schaefer, A.L.; MacLeod, E.K.; Zimenko, A.; Baker, D.; Greenberg, E.P. A Covariation analysis reveals elements of selectivity in quorum sensing systems. Elife 2021, 10, e69169. [Google Scholar] [CrossRef] [PubMed]
- Steindler, L.; Venturi, V. Detection of Quorum-sensing N-acyl homoserine lactone signal molecules by bacterial biosensors. FEMS Microbiol. Lett. 2007, 266, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deryabin, D.; Galadzhieva, A.; Kosyan, D.; Duskaev, G. Plant-derived inhibitors of AHL-mediated quorum sensing in bacteria: Modes of action. Int. J. Mol. Sci. 2019, 20, 5588. [Google Scholar] [CrossRef] [Green Version]
- Tobias, N.J.; Brehm, J.; Kresovic, D.; Brameyer, S.; Bode, H.B.; Heermann, R. New vocabulary for bacterial communication. ChemBioChem 2020, 21, 759–768. [Google Scholar] [CrossRef] [Green Version]
- Callahan, S.M.; Dunlap, P.V. LuxR- and acyl-homoserine-lactone-controlled Non-Lux genes define a quorum-sensing regulon in Vibrio fischeri. J. Bacteriol. 2000, 182, 2811–2822. [Google Scholar] [CrossRef] [Green Version]
- Bettenworth, V.; Steinfeld, B.; Duin, H.; Petersen, K.; Streit, W.R.; Bischofs, I.; Becker, A. Phenotypic heterogeneity in bacterial quorum sensing systems. J. Mol. Biol. 2019, 431, 4530–4546. [Google Scholar] [CrossRef]
- Kumar, M.; Saxena, M.; Saxena, A.K.; Nandi, S. Recent breakthroughs in various antimicrobial resistance induced quorum sensing biosynthetic pathway mediated targets and design of their inhibitors. Comb. Chem. High Throughput Screen. 2020, 23, 458–476. [Google Scholar] [CrossRef]
- Gould, T.A.; Herman, J.; Krank, J.; Murphy, R.C.; Churchill, M.E.A. Specificity of acyl-homoserine lactone synthases examined by mass spectrometry. J. Bacteriol. 2006, 188, 773–783. [Google Scholar] [CrossRef] [Green Version]
- Yi, L.; Li, J.; Liu, B.; Wang, Y. Advances in research on signal molecules regulating biofilms. World J. Microbiol. Biotechnol. 2019, 35, 130. [Google Scholar] [CrossRef]
- Burckhardt, R.M.; Escalante-Semerena, J.C. Small-molecule acetylation by GCN5-related N-acetyltransferases in bacteria. Microbiol. Mol. Biol. Rev. 2020, 84, e00090-19. [Google Scholar] [CrossRef] [PubMed]
- Tuon, F.F.; Dantas, L.R.; Suss, P.H.; Tasca Ribeiro, V.S. Pathogenesis of the Pseudomonas aeruginosa biofilm: A review. Pathogens 2022, 11, 300. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.W.; Khan, A.U. Updates on the pathogenicity status of Pseudomonas aeruginosa. Drug Discov. Today 2019, 24, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Holm, A.; Karlsson, T.; Vikström, E. Pseudomonas aeruginosa LasI/RhlI quorum sensing genes promote phagocytosis and aquaporin 9 redistribution to the leading and trailing regions in macrophages. Front. Microbiol. 2015, 6, 915. [Google Scholar] [CrossRef] [Green Version]
- Guła, G.; Dorotkiewicz-Jach, A.; Korzekwa, K.; Valvano, M.A.; Drulis-Kawa, Z. Complex signaling networks controlling dynamic molecular changes in Pseudomonas aeruginosa biofilm. Curr. Med. Chem. 2019, 26, 1979–1993. [Google Scholar] [CrossRef]
- Liu, L.; Zeng, X.; Zheng, J.; Zou, Y.; Qiu, S.; Dai, Y. AHL-mediated quorum sensing to regulate bacterial substance and energy metabolism: A review. Microbiol. Res. 2022, 262, 127102. [Google Scholar] [CrossRef]
- Lal, A.F.; Singh, S.; Franco, F.C.; Bhatia, S. Potential of polyphenols in curbing quorum sensing and biofilm formation in gram-negative pathogens. Asian Pac. J. Trop. Biomed. 2021, 11, 231. [Google Scholar] [CrossRef]
- González-Mula, A.; Lang, J.; Grandclément, C.; Naquin, D.; Ahmar, M.; Soulère, L.; Queneau, Y.; Dessaux, Y.; Faure, D. Lifestyle of the biotroph Agrobacterium tumefaciens in the ecological niche constructed on its host plant. New Phytol. 2018, 219, 350–362. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.; Su, S.; Farrand, S.K. Molecular basis of transcriptional antiactivation. TraM disrupts the TraR-DNA complex through stepwise interactions. J. Biol. Chem. 2007, 282, 19979–19991. [Google Scholar] [CrossRef] [Green Version]
- Fuqua, C.; Burbea, M.; Winans, S.C. Activity of the Agrobacterium Ti plasmid conjugal transfer regulator TraR Is inhibited by the product of the TraM gene. J. Bacteriol. 1995, 177, 1367–1373. [Google Scholar] [CrossRef]
- Gooderham, W.J.; Hancock, R.E.W. Regulation of virulence and antibiotic resistance by two-component regulatory systems in Pseudomonas aeruginosa. FEMS Microbiol. Rev. 2009, 33, 279–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Sun, D.; Zhu, J.; Liu, W. Two-component signal transduction systems: A major strategy for connecting input stimuli to biofilm formation. Front. Microbiol. 2019, 9, 3279. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, H.; Sivaneson, M.; Filloux, A. Key Two-component regulatory systems that control biofilm formation in Pseudomonas aeruginosa. Environ. Microbiol. 2011, 13, 1666–1681. [Google Scholar] [CrossRef] [PubMed]
- Okkotsu, Y.; Little, A.S.; Schurr, M.J. The Pseudomonas aeruginosa AlgZR two-component system coordinates multiple phenotypes. Front. Cell. Infect. Microbiol. 2014, 4, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abelyan, N.; Grabski, H.; Tiratsuyan, S. In silico screening of flavones and its derivatives as potential inhibitors of quorum-sensing regulator LasR of Pseudomonas aeruginosa. Mol. Biol. 2020, 54, 153–163. [Google Scholar] [CrossRef]
- Cabeen, M.T. Stationary phase-specific virulence factor overproduction by a LasR mutant of Pseudomonas aeruginosa. PLoS ONE 2014, 9, e88743. [Google Scholar] [CrossRef] [Green Version]
- Cocotl-Yañez, M.; Soto-Aceves, M.P.; González-Valdez, A.; Servín-González, L.; Soberón-Chávez, G. Virulence factors regulation by the quorum-sensing and Rsm systems in the marine strain Pseudomonas aeruginosa ID4365, a natural mutant in LasR. FEMS Microbiol. Lett. 2020, 367, fnaa092. [Google Scholar] [CrossRef]
- García-Reyes, S.; Soto-Aceves, M.P.; Cocotl-Yañez, M.; González-Valdez, A.; Servín-González, L.; Chávez, G.S. The Outlier Pseudomonas aeruginosa strain ATCC 9027 harbors a defective LasR quorum-sensing transcriptional regulator. FEMS Microbiol. Lett. 2020, 367, fnaa122. [Google Scholar] [CrossRef]
- He, Q.; Feng, Z.; Wang, Y.; Wang, K.; Zhang, K.; Kai, L.; Hao, X.; Yu, Z.; Chen, L.; Ge, Y. LasR might act as an intermediate in overproduction of phenazines in the absence of rpos in Pseudomonas aeruginosa. J. Microbiol. Biotechnol. 2019, 29, 1299–1309. [Google Scholar] [CrossRef]
- Hoffman, L.R.; Kulasekara, H.D.; Emerson, J.; Houston, L.S.; Burns, J.L.; Ramsey, B.W.; Miller, S.I. Pseudomonas aeruginosa LasR mutants are associated with cystic fibrosis lung disease progression. J. Cyst. Fibros. Off. J. Eur. Cyst. Fibros. Soc. 2009, 8, 66–70. [Google Scholar] [CrossRef]
- Manson, D.E.; O’Reilly, M.C.; Nyffeler, K.E.; Blackwell, H.E. Design, synthesis, and biochemical characterization of non-native antagonists of the Pseudomonas aeruginosa quorum sensing receptor LasR with nanomolar IC(50) values. ACS Infect. Dis. 2020, 6, 649–661. [Google Scholar] [CrossRef] [PubMed]
- McCready, A.R.; Paczkowski, J.E.; Henke, B.R.; Bassler, B.L. Structural determinants driving homoserine lactone ligand selection in the Pseudomonas aeruginosa LasR quorum-sensing receptor. Proc. Natl. Acad. Sci. USA 2019, 116, 245–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nain, Z.; Sayed, S.B.; Karim, M.M.; Islam, M.A.; Adhikari, U.K. Energy-optimized pharmacophore coupled virtual screening in the discovery of quorum sensing inhibitors of LasR protein of Pseudomonas aeruginosa. J. Biomol. Struct. Dyn. 2020, 38, 5374–5388. [Google Scholar] [CrossRef] [PubMed]
- O’Loughlin, C.T.; Miller, L.C.; Siryaporn, A.; Drescher, K.; Semmelhack, M.F.; Bassler, B.L. A Quorum-sensing inhibitor blocks Pseudomonas aeruginosa virulence and biofilm formation. Proc. Natl. Acad. Sci. USA 2013, 110, 17981–17986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadiq, S.; Rana, N.F.; Zahid, M.A.; Zargaham, M.K.; Tanweer, T.; Batool, A.; Naeem, A.; Nawaz, A.; Rizwan-Ur-Rehman; Muneer, Z.; et al. Virtual screening of FDA-approved drugs against LasR of Pseudomonas aeruginosa for antibiofilm potential. Molecules 2020, 25, 3723. [Google Scholar] [CrossRef]
- Abd El-Aleam, R.H.; George, R.F.; Georgey, H.H.; Abdel-Rahman, H.M. Bacterial virulence factors: A target for heterocyclic compounds to combat bacterial resistance. RSC Adv. 2021, 11, 36459–36482. [Google Scholar] [CrossRef]
- Rosales-Reyes, R.; Vargas-Roldán, S.Y.; Lezana-Fernández, J.L.; Santos-Preciado, J.I. Pseudomonas aeruginosa: Genetic adaptation, a strategy for its persistence in cystic fibrosis. Arch. Med. Res. 2021, 52, 357–361. [Google Scholar] [CrossRef]
- McCready, A.R.; Paczkowski, J.E.; Cong, J.-P.; Bassler, B.L. An autoinducer-independent RhlR quorum-sensing receptor enables analysis of RhlR regulation. PLoS Pathog. 2019, 15, e1007820. [Google Scholar] [CrossRef]
- Feltner, J.B.; Wolter, D.J.; Pope, C.E.; Groleau, M.-C.; Smalley, N.E.; Greenberg, E.P.; Mayer-Hamblett, N.; Burns, J.; Déziel, E.; Hoffman, L.R.; et al. LasR variant cystic fibrosis isolates reveal an adaptable quorum-sensing hierarchy in Pseudomonas aeruginosa. MBio 2016, 7, e01513-16. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Ahator, S.D.; Zhang, L.-H. Molecular mechanisms of phosphate stress activation of Pseudomonas aeruginosa quorum sensing systems. MSphere 2020, 5, e00119-20. [Google Scholar] [CrossRef]
- Rampioni, G.; Schuster, M.; Greenberg, E.P.; Bertani, I.; Grasso, M.; Venturi, V.; Zennaro, E.; Leoni, L. RsaL provides quorum sensing homeostasis and functions as a global regulator of gene expression in Pseudomonas aeruginosa. Mol. Microbiol. 2007, 66, 1557–1565. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.F.; Yang, C.; Zhang, Y.; Tao, S.N.; Mei, J.; Zhang, X.C.; Sun, Y.J.; Zhao, B.T. An innovative role for luteolin as a natural quorum sensing inhibitor in Pseudomonas aeruginosa. Life Sci. 2021, 274, 119325. [Google Scholar] [CrossRef] [PubMed]
- Berry, A.; Han, K.; Trouillon, J.; Robert-Genthon, M.; Ragno, M.; Lory, S.; Attrée, I.; Elsen, S. CAMP and Vfr control exolysin expression and cytotoxicity of Pseudomonas aeruginosa taxonomic outliers. J. Bacteriol. 2018, 200, e00135-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clay, M.E.; Hammond, J.H.; Zhong, F.; Chen, X.; Kowalski, C.H.; Lee, A.J.; Porter, M.S.; Hampton, T.H.; Greene, C.S.; Pletneva, E.V.; et al. Pseudomonas aeruginosa LasR mutant fitness in microoxia is supported by an anr-regulated oxygen-binding hemerythrin. Proc. Natl. Acad. Sci. USA 2020, 117, 3167–3173. [Google Scholar] [CrossRef] [PubMed]
- Maslova, E.; Eisaiankhongi, L.; Sjöberg, F.; McCarthy, R.R. Burns and biofilms: Priority pathogens and in vivo models. npj Biofilms Microbiomes 2021, 7, 73. [Google Scholar] [CrossRef]
- Choi, H.-Y.; Le, D.D.; Kim, W.-G. Curvularin Isolated from Phoma macrostoma Is an Antagonist of RhlR Quorum Sensing in Pseudomonas aeruginosa. Front. Microbiol. 2022, 13, 13882. [Google Scholar] [CrossRef]
- Ochsner, U.A.; Reiser, J. Autoinducer-mediated regulation of rhamnolipid biosurfactant synthesis in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1995, 92, 6424–6428. [Google Scholar] [CrossRef] [Green Version]
- Pearson, J.P.; Passador, L.; Iglewski, B.H.; Greenberg, E.P. A second N-acylhomoserine lactone signal produced by Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1995, 92, 1490–1494. [Google Scholar] [CrossRef] [Green Version]
- Pearson, J.P.; Pesci, E.C.; Iglewski, B.H. Roles of Pseudomonas aeruginosa Las and Rhl quorum-sensing systems in control of elastase and rhamnolipid biosynthesis genes. J. Bacteriol. 1997, 179, 5756–5767. [Google Scholar] [CrossRef] [Green Version]
- Li, L.-L.; Malone, J.E.; Iglewski, B.H. Regulation of the Pseudomonas aeruginosa quorum-sensing regulator VqsR. J. Bacteriol. 2007, 189, 4367–4374. [Google Scholar] [CrossRef]
- Medina, G.; Juárez, K.; Soberón-Chávez, G. The Pseudomonas aeruginosa RhlAB operon is not expressed during the logarithmic phase of growth even in the presence of its activator RhlR and the autoinducer N-butyryl-homoserine lactone. J. Bacteriol. 2003, 185, 377–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.A.K.S.; Rudden, M.; Smyth, T.J.; Dooley, J.S.G.; Marchant, R.; Banat, I.M. Natural quorum sensing inhibitors effectively downregulate gene expression of Pseudomonas aeruginosa virulence factors. Appl. Microbiol. Biotechnol. 2019, 103, 3521–3535. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, N.; Bagchi, A. Elucidation of the hetero-dimeric binding activity of LasR and RhlR proteins with the promoter dna and the role of a specific Phe residue during the biosynthesis of HCN synthase from opportunistic pathogen Pseudomonas aeruginosa. J. Mol. Model. 2021, 27, 76. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, S.; Bhatia, S.; Singh, S.; Franco, F.J. Growing emergence of drug-resistant Pseudomonas aeruginosa and attenuation of its virulence using quorum sensing inhibitors: A critical review. Iran. J. Basic Med. Sci. 2021, 24, 699–719. [Google Scholar] [CrossRef] [PubMed]
- Ledgham, F.; Soscia, C.; Chakrabarty, A.; Lazdunski, A.; Foglino, M. Global regulation in Pseudomonas aeruginosa: The regulatory protein AlgR2 (AlgQ) acts as a modulator of quorum sensing. Res. Microbiol. 2003, 154, 207–213. [Google Scholar] [CrossRef]
- Chugani, S.A.; Whiteley, M.; Lee, K.M.; D’Argenio, D.; Manoil, C.; Greenberg, E.P. QscR, a modulator of quorum-sensing signal synthesis and virulence in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2001, 98, 2752–2757. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Wang, M.; Smalley, N.E.; Kostylev, M.; Schaefer, A.L.; Greenberg, E.P.; Dandekar, A.A.; Xu, F. Modulation of Pseudomonas aeruginosa quorum sensing by glutathione. J. Bacteriol. 2019, 201, e00685-18. [Google Scholar] [CrossRef] [Green Version]
- Wysoczynski-Horita, C.L.; Boursier, M.E.; Hill, R.; Hansen, K.; Blackwell, H.E.; Churchill, M.E.A. Mechanism of agonism and antagonism of the Pseudomonas aeruginosa quorum sensing regulator QscR with non-native ligands. Mol. Microbiol. 2018, 108, 240–257. [Google Scholar] [CrossRef] [Green Version]
- Barton, I.S.; Eagan, J.L.; Nieves-Otero, P.A.; Reynolds, I.P.; Platt, T.G.; Fuqua, C. Co-dependent and interdigitated: Dual quorum sensing systems regulate conjugative transfer of the Ti plasmid and the at megaplasmid in Agrobacterium tumefaciens 15955. Front. Microbiol. 2021, 11, 605896. [Google Scholar] [CrossRef]
- Faure, D.; Lang, J. Functions and regulation of quorum-sensing in Agrobacterium tumefaciens. Front. Plant Sci. 2014, 5, 14. [Google Scholar] [CrossRef]
- Luo, Z.Q.; Farrand, S.K. Signal-dependent DNA binding and functional domains of the quorum-sensing activator TraR as identified by repressor activity. Proc. Natl. Acad. Sci. USA 1999, 96, 9009–9014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, C.E.; Winans, S.C. Identification of amino acid residues of the Agrobacterium tumefaciens quorum-sensing regulator TraR that are critical for positive control of transcription. Mol. Microbiol. 2005, 55, 1473–1486. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, M.E.; Asenstorfer, R.E.; Tate, M.E.; Farrand, S.K. Quorum-dependent transfer of the opine-catabolic plasmid PAoF64/95 is regulated by a novel mechanism involving inhibition of the TraR antiactivator TraM. Microbiologyopen 2019, 8, e00625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, C.E.; Winans, S.C. The quorum-sensing transcription factor TraR decodes its DNA binding site by direct contacts with DNA bases and by detection of DNA flexibility. Mol. Microbiol. 2007, 64, 245–256. [Google Scholar] [CrossRef]
- Qin, Y.; Luo, Z.-Q.; Farrand, S.K. Domains formed within the N-terminal region of the quorum-sensing activator TraR are required for transcriptional activation and direct interaction with RpoA from Agrobacterium. J. Biol. Chem. 2004, 279, 40844–40851. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Winans, S.C. Autoinducer binding by the quorum-sensing regulator TraR increases affinity for target promoters in vitro and decreases TraR turnover rates in whole cells. Proc. Natl. Acad. Sci. USA 1999, 96, 4832–4837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappas, K.M.; Winans, S.C. A luxR-type regulator from Agrobacterium tumefaciens elevates Ti plasmid copy number by activating transcription of plasmid replication genes. Mol. Microbiol. 2003, 48, 1059–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Y.; Luo, Z.Q.; Smyth, A.J.; Gao, P.; Beck von Bodman, S.; Farrand, S.K. Quorum-sensing signal binding results in dimerization of TraR and its release from membranes into the cytoplasm. EMBO J. 2000, 19, 5212–5221. [Google Scholar] [CrossRef]
- Vreede, J.; Hellingwerf, K.J.; Crielaard, W. TraR auto-inducer enhances protein backbone fluctuations in DNA binding domain. FEBS Lett. 2008, 582, 805–809. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.; Winans, S.C. TraA, TraC and TraD autorepress two divergent quorum-regulated promoters near the transfer origin of the Ti plasmid of Agrobacterium tumefaciens. Mol. Microbiol. 2007, 63, 1769–1782. [Google Scholar] [CrossRef]
- Li, P.L.; Everhart, D.M.; Farrand, S.K. Genetic and sequence analysis of the PTiC58 Trb locus, encoding a mating-pair formation system related to members of the type IV secretion family. J. Bacteriol. 1998, 180, 6164–6172. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Pinto, U.M.; Winans, S.C. Transsexuality in the rhizosphere: Quorum sensing reversibly converts Agrobacterium tumefaciens from phenotypically female to male. J. Bacteriol. 2009, 191, 3375–3383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, Y.; Winans, S.C. The Chaperone GroESL Enhances the accumulation of soluble, active TraR protein, a quorum-sensing transcription factor from Agrobacterium tumefaciens. J. Bacteriol. 2009, 191, 3706–3711. [Google Scholar] [CrossRef] [Green Version]
- Swiderska, A.; Berndtson, A.K.; Cha, M.R.; Li, L.; Beaudoin, G.M., 3rd; Zhu, J.; Fuqua, C. Inhibition of the Agrobacterium tumefaciens TraR quorum-sensing regulator: Interactions with the TraM anti-activator. J. Biol. Chem. 2001, 276, 49449–49458. [Google Scholar] [CrossRef] [Green Version]
- Vannini, A.; Volpari, C.; Gargioli, C.; Muraglia, E.; Cortese, R.; De Francesco, R.; Neddermann, P.; Marco, S. Di the crystal structure of the quorum sensing protein TraR bound to its autoinducer and target DNA. EMBO J. 2002, 21, 4393–4401. [Google Scholar] [CrossRef] [PubMed]
- Stauff, D.L.; Bassler, B.L. Quorum sensing in Chromobacterium violaceum: DNA recognition and gene regulation by the CviR receptor. J. Bacteriol. 2011, 193, 3871–3878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durán, N.; Menck, C.F. Chromobacterium violaceum: A review of pharmacological and industrial perspectives. Crit. Rev. Microbiol. 2001, 27, 201–222. [Google Scholar] [CrossRef] [PubMed]
- Morohoshi, T.; Fukamachi, K.; Kato, M.; Kato, N.; Ikeda, T. Regulation of the violacein biosynthetic gene cluster by acylhomoserine lactone-mediated quorum sensing in Chromobacterium violaceum ATCC 12472. Biosci. Biotechnol. Biochem. 2010, 74, 2116–2119. [Google Scholar] [CrossRef]
- Chernin, L.S.; Winson, M.K.; Thompson, J.M.; Haran, S.; Bycroft, B.W.; Chet, I.; Williams, P.; Stewart, G.S. Chitinolytic activity in Chromobacterium violaceum: Substrate analysis and regulation by quorum sensing. J. Bacteriol. 1998, 180, 4435–4441. [Google Scholar] [CrossRef] [Green Version]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef]
- Kumar, L.; Brenner, N.; Brice, J.; Klein-Seetharaman, J.; Sarkar, S.K. Cephalosporins interfere with quorum sensing and improve the ability of Caenorhabditis elegans to survive Pseudomonas aeruginosa infection. Front. Microbiol. 2021, 12, 598498. [Google Scholar] [CrossRef] [PubMed]
- Vasavi, H.S.; Arun, A.B.; Rekha, P.-D. Anti-quorum sensing activity of Psidium guajava L. flavonoids against Chromobacterium violaceum and Pseudomonas aeruginosa PAO1. Microbiol. Immunol. 2014, 58, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Moradi, F.; Hadi, N.; Bazargani, A. Evaluation of quorum-sensing inhibitory effects of extracts of three traditional medicine plants with known antibacterial properties. New Microbes New Infect. 2020, 38, 100769. [Google Scholar] [CrossRef] [PubMed]
- Deryabin, D.G.; Tolmacheva, A.A. Antibacterial and anti-quorum sensing molecular composition derived from Quercus cortex (Oak Bark) extract. Molecules 2015, 20, 17093–17108. [Google Scholar] [CrossRef] [Green Version]
- Ravichandran, V.; Zhong, L.; Wang, H.; Yu, G.; Zhang, Y.; Li, A. Virtual screening and biomolecular interactions of CviR-based quorum sensing inhibitors against Chromobacterium violaceum. Front. Cell. Infect. Microbiol. 2018, 8, 298. [Google Scholar] [CrossRef]
- Chen, G.; Swem, L.R.; Swem, D.L.; Stauff, D.L.; O’Loughlin, C.T.; Jeffrey, P.D.; Bassler, B.L.; Hughson, F.M. A strategy for antagonizing quorum sensing. Mol. Cell 2011, 42, 199–209. [Google Scholar] [CrossRef] [Green Version]
- Michael, B.; Smith, J.N.; Swift, S.; Heffron, F.; Ahmer, B.M. SdiA of Salmonella enterica is a LuxR homolog that detects mixed microbial communities. J. Bacteriol. 2001, 183, 5733–5742. [Google Scholar] [CrossRef] [Green Version]
- Joseleau-Petit, D.; Vinella, D.; D’Ari, R. Metabolic Alarms and Cell Division in Escherichia Coli. J. Bacteriol. 1999, 181, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.D.; de Boer, P.A.; Rothfield, L.I. A factor that positively regulates cell division by activating transcription of the major cluster of essential cell division genes of Escherichia coli. EMBO J. 1991, 10, 3363–3372. [Google Scholar] [CrossRef]
- Yao, Y.; Martinez-Yamout, M.A.; Dickerson, T.J.; Brogan, A.P.; Wright, P.E.; Dyson, H.J. Structure of the Escherichia coli quorum sensing protein SdiA: Activation of the folding switch by acyl homoserine lactones. J. Mol. Biol. 2006, 355, 262–273. [Google Scholar] [CrossRef]
- Styles, M.J.; Early, S.A.; Tucholski, T.; West, K.H.J.; Ge, Y.; Blackwell, H.E. Chemical control of quorum sensing in E. Coli: Identification of small molecule modulators of SdiA and mechanistic characterization of a covalent inhibitor. ACS Infect. Dis. 2020, 6, 3092–3103. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Y.; Nguyen, N.X.; Rogers, J.L.; Liao, J.; MacMillan, J.B.; Jiang, Y.; Sperandio, V. Structural and mechanistic roles of novel chemical ligands on the SdiA quorum-sensing transcription regulator. MBio 2015, 6, e02429-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanamaru, K.; Kanamaru, K.; Tatsuno, I.; Tobe, T.; Sasakawa, C. SdiA, an Escherichia Coli homologue of quorum-sensing regulators, controls the expression of virulence factors in enterohaemorrhagic Escherichia coli O157:H7. Mol. Microbiol. 2000, 38, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.K.; Bearson, S.M.D.; Bearson, B.L. Evaluation of the Eeffects of SdiA, a LuxR homologue, on adherence and motility of Escherichia Coli O157:H7. Microbiology 2010, 156, 1303–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culler, H.F.; Couto, S.C.F.; Higa, J.S.; Ruiz, R.M.; Yang, M.J.; Bueris, V.; Franzolin, M.R.; Sircili, M.P. Role of SdiA on biofilm formation by atypical enteropathogenic Escherichia coli. Genes 2018, 9, 253. [Google Scholar] [CrossRef] [Green Version]
- Tavío, M.M.; Aquili, V.D.; Poveda, J.B.; Antunes, N.T.; Sánchez-Céspedes, J.; Vila, J. Quorum-sensing regulator SdiA and MarA overexpression is involved in in vitro-selected multidrug resistance of Escherichia coli. J. Antimicrob. Chemother. 2010, 65, 1178–1186. [Google Scholar] [CrossRef]
- Sitnikov, D.M.; Schineller, J.B.; Baldwin, T.O. Control of cell division in Escherichia coli: Regulation of transcription of FtsQA involves both RpoS and SdiA-mediated autoinduction. Proc. Natl. Acad. Sci. USA 1996, 93, 336–341. [Google Scholar] [CrossRef] [Green Version]
- Sperandio, V. SdiA sensing of acyl-homoserine lactones by Enterohemorrhagic E. coli (EHEC) serotype O157:H7 in the bovine rumen. Gut Microbes 2010, 1, 432–435. [Google Scholar] [CrossRef] [Green Version]
- Grabski, H.; Ginosyan, S.; Tiratsuyan, S. Molecular simulations and markov state modeling reveal inactive form of quorum sensing regulator SdiA of Escherichia coli. IEEE/ACM Trans. Comput. Biol. Bioinforma. 2021, 18, 2835–2840. [Google Scholar] [CrossRef]
- Miller, C.; Gilmore, J. Detection of quorum-sensing molecules for pathogenic molecules using cell-based and cell-free biosensors. Antibiotics 2020, 9, 259. [Google Scholar] [CrossRef]
- Defoirdt, T. Quorum-sensing systems as targets for antivirulence therapy. Trends Microbiol. 2018, 26, 313–328. [Google Scholar] [CrossRef] [PubMed]
- John, J.; Saranathan, R.; Adigopula, L.N.; Thamodharan, V.; Singh, S.P.; Lakshmi, T.P.; CharanTej, M.A.; Rao, R.S.; Krishna, R.; Rao, H.S.P.; et al. The quorum sensing molecule N-acyl homoserine lactone produced by Acinetobacter baumannii displays antibacterial and anticancer properties. Biofouling 2016, 32, 1029–1047. [Google Scholar] [CrossRef] [PubMed]
- Paluch, E.; Rewak-Soroczyńska, J.; Jędrusik, I.; Mazurkiewicz, E.; Jermakow, K. Prevention of biofilm formation by quorum quenching. Appl. Microbiol. Biotechnol. 2020, 104, 1871–1881. [Google Scholar] [CrossRef] [Green Version]
- Kalia, V.C.; Patel, S.K.S.; Kang, Y.C.; Lee, J.-K. Quorum sensing inhibitors as antipathogens: Biotechnological applications. Biotechnol. Adv. 2019, 37, 68–90. [Google Scholar] [CrossRef] [PubMed]
- Parasuraman, P.; Devadatha, B.; Sarma, V.V.; Ranganathan, S.; Ampasala, D.R.; Reddy, D.; Kumavath, R.; Kim, I.-W.; Patel, S.K.S.; Kalia, V.C.; et al. Inhibition of microbial quorum sensing mediated virulence factors by Pestalotiopsis sydowiana. J. Microbiol. Biotechnol. 2020, 30, 571–582. [Google Scholar] [CrossRef]
- Kumar, L.; Chhibber, S.; Kumar, R.; Kumar, M.; Harjai, K. Zingerone silences quorum sensing and attenuates virulence of Pseudomonas aeruginosa. Fitoterapia 2015, 102, 84–95. [Google Scholar] [CrossRef]
- Li, L.; Hooi, D.; Chhabra, S.R.; Pritchard, D.; Shaw, P.E. Bacterial N-acylhomoserine lactone-induced apoptosis in breast carcinoma cells correlated with down-modulation of STAT3. Oncogene 2004, 23, 4894–4902. [Google Scholar] [CrossRef] [Green Version]
- Khajanchi, B.K.; Kirtley, M.L.; Brackman, S.M.; Chopra, A.K. Immunomodulatory and protective roles of quorum-sensing signaling molecules N-acyl homoserine lactones during infection of mice with Aeromonas hydrophila. Infect. Immun. 2011, 79, 2646–2657. [Google Scholar] [CrossRef] [Green Version]
- Miyairi, S.; Tateda, K.; Fuse, E.T.; Ueda, C.; Saito, H.; Takabatake, T.; Ishii, Y.; Horikawa, M.; Ishiguro, M.; Standiford, T.J.; et al. Immunization with 3-oxododecanoyl-L-homoserine lactone-protein conjugate protects mice from lethal Pseudomonas aeruginosa lung infection. J. Med. Microbiol. 2006, 55, 1381–1387. [Google Scholar] [CrossRef]
- Vasudevan, S.; Srinivasan, P.; Rayappan, J.B.B.; Solomon, A.P. A photoluminescence biosensor for the detection of N-acyl homoserine lactone using cysteamine functionalized ZnO nanoparticles for the early diagnosis of urinary tract infections. J. Mater. Chem. B 2020, 8, 4228–4236. [Google Scholar] [CrossRef]
- Bodini, S.F.; Manfredini, S.; Epp, M.; Valentini, S.; Santori, F. Quorum sensing inhibition activity of garlic extract and p-coumaric acid. Lett. Appl. Microbiol. 2009, 49, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Miari, M.; Rasheed, S.S.; Haidar-Ahmad, N.; Abou Fayad, A.; Matar, G.M. Evaluating ginger extract, wild blueberry extract, and polysorbates (PS20, PS80) on Pseudomonas aeruginosa biofilm formation. J. Infect. Dev. Ctries. 2018, 12, 12S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maisuria, V.B.; Los Santos, Y.L.; Tufenkji, N.; Déziel, E. Cranberry-derived proanthocyanidins impair virulence and inhibit quorum sensing of Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 30169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shastry, R.P.; Ghate, S.D.; Sukesh Kumar, B.; Srinath, B.S.; Kumar, V. Vanillin derivative inhibits quorum sensing and biofilm formation in Pseudomonas aeruginosa: A study in a Caenorhabditis elegans infection model. Nat. Prod. Res. 2022, 36, 1610–1615. [Google Scholar] [CrossRef]
- Abdulrahman, H.; Misba, L.; Ahmad, S.; Khan, A.U. Curcumin induced photodynamic therapy mediated suppression of quorum sensing pathway of Pseudomonas aeruginosa: An approach to inhibit biofilm in vitro. Photodiagnosis Photodyn. Ther. 2020, 30, 101645. [Google Scholar] [CrossRef]
- Dulla, G.F.J.; Krasileva, K.V.; Lindow, S.E. Interference of quorum sensing in Pseudomonas syringae by bacterial epiphytes that limit iron availability. Environ. Microbiol. 2010, 12, 1762–1774. [Google Scholar] [CrossRef]
- Dulla, G.F.J.; Lindow, S.E. Acyl-homoserine lactone-mediated cross talk among epiphytic bacteria modulates behavior of Pseudomonas syringae on leaves. ISME J. 2009, 3, 825–834. [Google Scholar] [CrossRef]
- Hartmann, A.; Rothballer, M.; Hense, B.A.; Schröder, P. Bacterial quorum sensing compounds are important modulators of microbe-plant interactions. Front. Plant Sci. 2014, 5, 131. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.-Z.; Morohoshi, T.; Ikenoya, M.; Someya, N.; Ikeda, T. AiiM, a novel class of N-acylhomoserine lactonase from the leaf-associated bacterium Microbacterium testaceum. Appl. Environ. Microbiol. 2010, 76, 2524–2530. [Google Scholar] [CrossRef] [Green Version]
- De Celis, M.; Serrano-Aguirre, L.; Belda, I.; Liébana-García, R.; Arroyo, M.; Marquina, D.; de la Mata, I.; Santos, A. Acylase enzymes disrupting quorum sensing alter the transcriptome and phenotype of Pseudomonas aeruginosa, and the composition of bacterial biofilms from wastewater treatment plants. Sci. Total Environ. 2021, 799, 149401. [Google Scholar] [CrossRef]
- Rémy, B.; Plener, L.; Decloquement, P.; Armstrong, N.; Elias, M.; Daudé, D.; Chabrière, É. Lactonase specificity is key to quorum quenching in Pseudomonas aeruginosa. Front. Microbiol. 2020, 11, 762. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.H.; Xu, J.L.; Li, X.Z.; Zhang, L.H. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc. Natl. Acad. Sci. USA 2000, 97, 3526–3531. [Google Scholar] [CrossRef] [PubMed]
- Teplitski, M.; Mathesius, U.; Rumbaugh, K.P. Perception and degradation of N-acyl homoserine lactone quorum sensing signals by mammalian and plant cells. Chem. Rev. 2011, 111, 100–116. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, A.; Rothballer, M. Role of quorum sensing signals of rhizobacteria for plant growth promotion. In Rhizotrophs: Plant Growth Promotion to Bioremediation; Springer: Berlin/Heidelberg, Germany, 2017; pp. 205–217. [Google Scholar] [CrossRef]
- Mathesius, U.; Mulders, S.; Gao, M.; Teplitski, M.; Caetano-Anollés, G.; Rolfe, B.G.; Bauer, W.D. Extensive and specific responses of a Eukaryote to bacterial quorum-sensing signals. Proc. Natl. Acad. Sci. USA 2003, 100, 1444–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuhegger, R.; Ihring, A.; Gantner, S.; Bahnweg, G.; Knappe, C.; Vogg, G.; Hutzler, P.; Schmid, M.; Van Breusegem, F.; Eberl, L.E.O. Induction of systemic resistance in tomato by N-acyl-L-homoserine lactone-producing rhizosphere bacteria. Plant. Cell Environ. 2006, 29, 909–918. [Google Scholar] [CrossRef]
- Müller, H.; Westendorf, C.; Leitner, E.; Chernin, L.; Riedel, K.; Schmidt, S.; Eberl, L.; Berg, G. Quorum-sensing effects in the antagonistic rhizosphere bacterium Serratia plymuthica HRO-C48. FEMS Microbiol. Ecol. 2009, 67, 468–478. [Google Scholar] [CrossRef] [Green Version]
- Von Rad, U.; Klein, I.; Dobrev, P.I.; Kottova, J.; Zazimalova, E.; Fekete, A.; Hartmann, A.; Schmitt-Kopplin, P.; Durner, J. Response of Arabidopsis thaliana to N-hexanoyl-DL-homoserine-lactone, a bacterial quorum sensing molecule produced in the rhizosphere. Planta 2008, 229, 73–85. [Google Scholar] [CrossRef]
- Lade, H.; Paul, D.; Kweon, J.H. Quorum quenching mediated approaches for control of membrane biofouling. Int. J. Biol. Sci. 2014, 10, 550–565. [Google Scholar] [CrossRef]
- Ponnusamy, K.; Kappachery, S.; Thekeettle, M.; Song, J.H.; Kweon, J.H. Anti-biofouling property of vanillin on Aeromonas hydrophila initial biofilm on various membrane surfaces. World J. Microbiol. Biotechnol. 2013, 29, 1695–1703. [Google Scholar] [CrossRef]
- Yeon, K.-M.; Lee, C.-H.; Kim, J. Magnetic enzyme carrier for effective biofouling control in the membrane bioreactor based on enzymatic quorum quenching. Environ. Sci. Technol. 2009, 43, 7403–7409. [Google Scholar] [CrossRef]
- Uroz, S.; Chhabra, S.R.; Cámara, M.; Williams, P.; Oger, P.; Dessaux, Y. N-acylhomoserine lactone quorum-sensing molecules are modified and degraded by Rhodococcus erythropolis W2 by both amidolytic and novel oxidoreductase activities. Microbiology 2005, 151, 3313–3322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, Y.-C.; Zhong, J.-J. N-acylated homoserine lactone production and involvement in the biodegradation of aromatics by an environmental isolate of Pseudomonas aeruginosa. Process Biochem. 2010, 45, 1944–1948. [Google Scholar] [CrossRef]
- Huang, Y.; Zeng, Y.; Yu, Z.; Zhang, J.; Feng, H.; Lin, X. In silico and experimental methods revealed highly diverse bacteria with quorum sensing and aromatics biodegradation systems—A potential broad application on bioremediation. Bioresour. Technol. 2013, 148, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.-S.; Park, W. Contribution of quorum-sensing system to hexadecane degradation and biofilm formation in Acinetobacter Sp. strain DR1. J. Appl. Microbiol. 2010, 109, 1650–1659. [Google Scholar] [CrossRef]
- Tomei, M.C.; Mosca Angelucci, D.; Clagnan, E.; Brusetti, L. Anaerobic biodegradation of phenol in wastewater treatment: Achievements and limits. Appl. Microbiol. Biotechnol. 2021, 105, 2195–2224. [Google Scholar] [CrossRef]
- Rahman, M.M.; Rahaman, M.S.; Islam, M.R.; Rahman, F.; Mithi, F.M.; Alqahtani, T.; Almikhlafi, M.A.; Alghamdi, S.Q.; Alruwaili, A.S.; Hossain, M.S.; et al. Role of phenolic compounds in human disease: Current knowledge and future prospects. Molecules 2021, 27, 233. [Google Scholar] [CrossRef]
- Hasan, S.A.; Jabeen, S. Degradation kinetics and pathway of phenol by Pseudomonas and Bacillus species. Biotechnol. Biotechnol. Equip. 2015, 29, 45–53. [Google Scholar] [CrossRef]
- Nde, C.W.; Jang, H.-J.; Toghrol, F.; Bentley, W.E. Toxicogenomic response of Pseudomonas aeruginosa to ortho-phenylphenol. BMC Genomics 2008, 9, 473. [Google Scholar] [CrossRef] [Green Version]
- Santos, P.M.; Benndorf, D.; Sá-Correia, I. Insights into Pseudomonas putida KT2440 response to phenol-induced stress by quantitative proteomics. Proteomics 2004, 4, 2640–2652. [Google Scholar] [CrossRef]
- Valle, A.; Bailey, M.J.; Whiteley, A.S.; Manefield, M. N-Acyl-L-homoserine lactones (AHLs) affect microbial community composition and function in activated sludge. Environ. Microbiol. 2004, 6, 424–433. [Google Scholar] [CrossRef]
- Cui, X.; Ruan, X.; Yin, J.; Wang, M.; Li, N.; Shen, D. Regulation of Las and Rhl quorum sensing on aerobic denitrification in Pseudomonas aeruginosa PAO1. Curr. Microbiol. 2021, 78, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Noh, J.; Park, W. Physiological and metabolic responses for hexadecane degradation in Acinetobacter oleivorans DR1. J. Microbiol. 2011, 49, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Chugani, S.; Greenberg, E.P. LuxR homolog-independent gene regulation by acyl-homoserine lactones in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2010, 107, 10673–10678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyofuku, M.; Nomura, N.; Kuno, E.; Tashiro, Y.; Nakajima, T.; Uchiyama, H. Influence of the Pseudomonas quinolone signal on denitrification in Pseudomonas aeruginosa. J. Bacteriol. 2008, 190, 7947–7956. [Google Scholar] [CrossRef] [Green Version]
- Silagyi, K.; Kim, S.-H.; Lo, Y.M.; Wei, C. Production of biofilm and quorum sensing by Escherichia coli O157:H7 and its transfer from contact surfaces to meat, poultry, ready-to-eat deli, and produce products. Food Microbiol. 2009, 26, 514–519. [Google Scholar] [CrossRef]
- Yadav, M.; Shukla, P. Efficient engineered probiotics using synthetic biology approaches: A review. Biotechnol. Appl. Biochem. 2020, 67, 22–29. [Google Scholar] [CrossRef]
- Meena, H.; Mishra, R.; Ranganathan, S.; Sarma, V.V.; Ampasala, D.R.; Kalia, V.C.; Lee, J.-K.; Siddhardha, B. Phomopsis tersa as inhibitor of quorum sensing system and biofilm forming ability of Pseudomonas aeruginosa. Indian J. Microbiol. 2020, 60, 70–77. [Google Scholar] [CrossRef]
- Anderson, J.C.; Clarke, E.J.; Arkin, A.P.; Voigt, C.A. Environmentally controlled invasion of cancer cells by engineered bacteria. J. Mol. Biol. 2006, 355, 619–627. [Google Scholar] [CrossRef]
- Wynendaele, E.; Verbeke, F.; D’Hondt, M.; Hendrix, A.; Van De Wiele, C.; Burvenich, C.; Peremans, K.; De Wever, O.; Bracke, M.; De Spiegeleer, B. Crosstalk between the microbiome and cancer cells by quorum sensing peptides. Peptides 2015, 64, 40–48. [Google Scholar] [CrossRef]
- Kalia, V.C.; Patel, S.K.S.; Cho, B.-K.; Wood, T.K.; Lee, J.-K. Emerging applications of bacteria as antitumor agents. Sem. Cancer Biol. 2021, 86, 1014–1025. [Google Scholar] [CrossRef]
- Gambello, M.J.; Kaye, S.; Iglewski, B.H. LasR of Pseudomonas aeruginosa is a transcriptional activator of the alkaline protease gene (Apr) and an enhancer of exotoxin a expression. Infect. Immun. 1993, 61, 1180–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rust, L.; Pesci, E.C.; Iglewski, B.H. Analysis of the Pseudomonas aeruginosa elastase (LasB) regulatory region. J. Bacteriol. 1996, 178, 1134–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateu-Borrás, M.; Zamorano, L.; González-Alsina, A.; Sánchez-Diener, I.; Doménech-Sánchez, A.; Oliver, A.; Albertí, S. Molecular analysis of the contribution of alkaline protease A and elastase B to the virulence of Pseudomonas aeruginosa bloodstream infections. Front. Cell. Infect. Microbiol. 2022, 11, 816356. [Google Scholar] [CrossRef] [PubMed]
- Seed, P.C.; Passador, L.; Iglewski, B.H. Activation of the Pseudomonas aeruginosa LasI gene by LasR and the Pseudomonas autoinducer PAI: An autoinduction regulatory hierarchy. J. Bacteriol. 1995, 177, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simanek, K.A.; Taylor, I.R.; Richael, E.K.; Lasek-Nesselquist, E.; Bassler, B.L.; Paczkowski, J.E. The PqsE-RhlR interaction regulates RhlR DNA binding to control virulence factor production in Pseudomonas aeruginosa. Microbiol. Spectr. 2022, 10, e0210821. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, M.; Lee, K.M.; Greenberg, E.P. Identification of genes controlled by quorum sensing in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1999, 96, 13904–13909. [Google Scholar] [CrossRef] [Green Version]
- Lequette, Y.; Lee, J.-H.; Ledgham, F.; Lazdunski, A.; Greenberg, E.P. A distinct QscR regulon in the Pseudomonas aeruginosa quorum-sensing circuit. J. Bacteriol. 2006, 188, 3365–3370. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.N.; Ahmer, B.M.M. Detection of other microbial species by Salmonella: Expression of the SdiA regulon. J. Bacteriol. 2003, 185, 1357–1366. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Function | Active-Site Residue |
|---|---|---|
| FeeM | Acyl-ACP-dependent amino acid acyltransferase | Ala168, Try32, Tyr 163, Val166, Pro169, Tyr27, Ala170, Asn142, Phe38, Ile141, Ser140, Gln97, Val96, Ile139, Glu94, Tyr149, Val95, Phe154, Leu175, Phe122, Val125, and Leu126 |
| LasI | 3-oxo-C12-HSL production | Ile168, Trp33, Glu171, Arg172, Ala173, Phe27, Ile43, Met79, Thr145, Val143, Thr144, Arg104, Ser103, Thr142, Phe105, Glu101, Leu102, met152, Leu140, Met125, Leu128, and Ala129 |
| RhlI | C4-HSL production | Ile171, Trp35, Ile175, Ala176, Ile177, Phe29, Ile177, Asp148, Met79, Arg45, Thr147, Thr145, Arg104, Ser103, Phe105, Glu101, Ser102, Leu155, Ile143, Phe128, Trp160, Ile131, and Ile132 |
| CviI | C6-HSL production | Val154, Leu155, Ser156, Glu127, Val126, Val125, Arg86, Ser85, Phe124, Phe87, Glu83, Ile84, Met122, Leu134, Tyr107, Leu110, Phe139, and Trp111 |
| TraI | 3-oxo-C8-HSL detection | Val163, Glu166, Trp34, Thr167, Leu168, Thr140, Val138, Thr139, Ala137, Phe28, Leu80, Phe47, Arg104, Ser103, Tyr105, Glu101, Leu102, Val135, Val152, Phe120, Ser123, and Leu124 |
| LasR | 3-oxo-C12-HSL detection | Leu36, Gly38, Leu38, Leu38, Tyr47, Ala50, Ile52, Arg61, Tyr64, Ala70, Thr75, Val76, Cys79, Trp88, Tyr93, Phe101, Ala105, Leu110, Thr115, Leu125, Gly126, and Ala127. |
| RhlR | C4-HSL detection | Arg48, Glu59, Tyr72, Ala83, Ile84, Trp96, Ala111, Leu116, and Thr121 |
| QscR | 3-oxo-C12-HSL detection | Phe39, Gly40, Ala41, Arg42, Tyr52, Phe54, Tyr66, Thr72, Ile77, Val78, and Gly81 Leu82, Trp90, Phe101, Trp102, Ala105, Ile110, Pro117, Ile125, Ser126, Met127, and Ser129 |
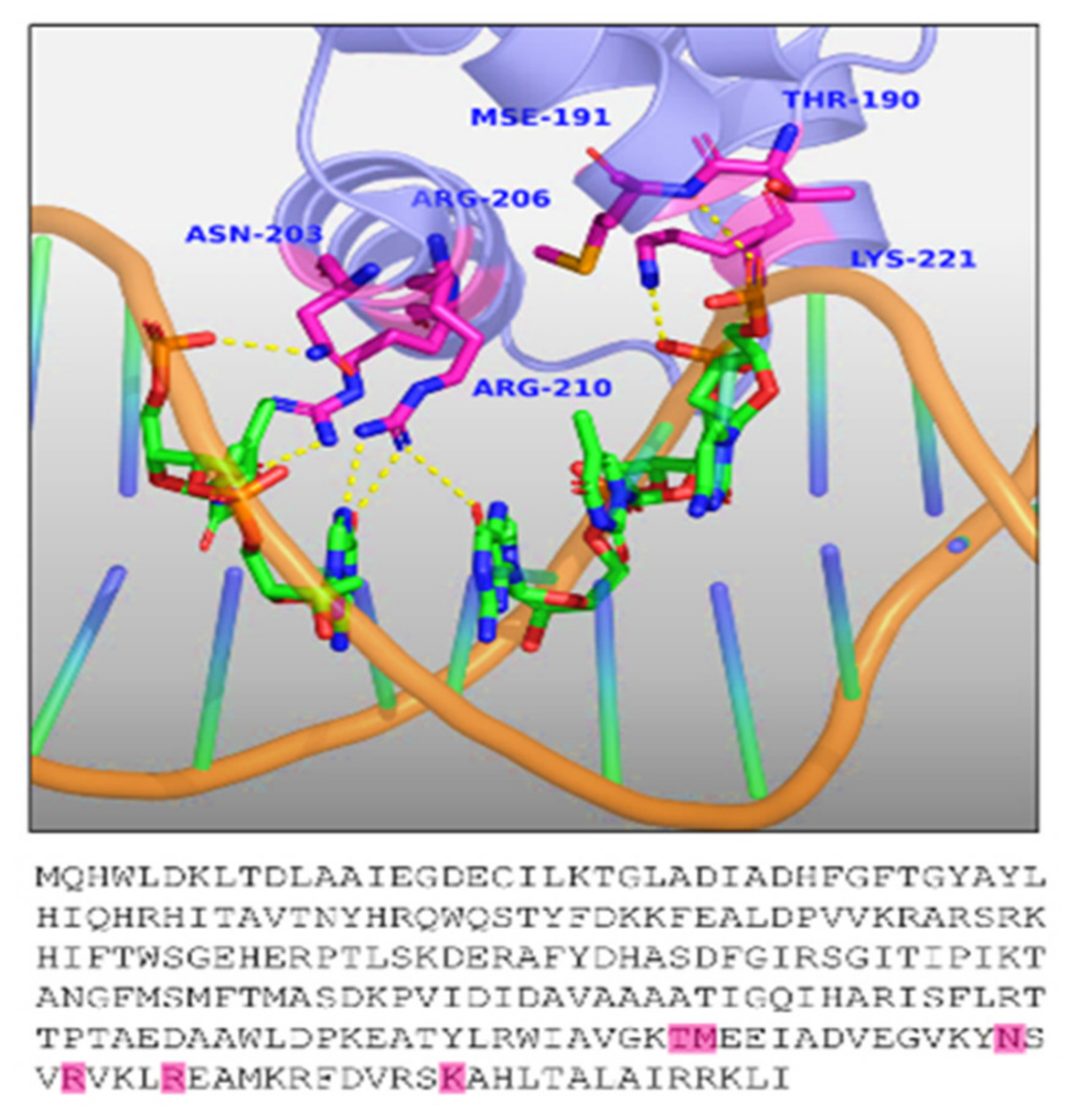
| TraR | 3-oxo-C12-HSL detection | Ala38, Leu40, Ala49, Thr51, Gln58, Tyr61, Phe62, Val73, Val74, Trp85, Phe101, Tyr102, Ala105, Ile110, Thr115, Mse127, and Ala168 |
| CviR | C6-HSL detection | Leu57, Val75, Leu85, Tyr88, Ile99, Leu100, Trp111, Phe115, Phe126, Ala130, Met135, Thr140, and Ile153 |
| SdiA | 3-oxo-C12-HSL detection | Cys45, Arg47, Val57, Ala58, Phe59, Tyr63, Tyr71, Leu77, Val72, Leu77, Trp95, Phe100, Leu106, Ala110, Leu115, Leu130, and Phe132 |
| QS Regulator | Phenotype/ Virulence Factor | Genetic Marker | Function | References |
|---|---|---|---|---|
| LasI/R | Protease | lasA | Epithelial barrier disruption, the adaptation and spread of infection, and immune evasion | [192] |
| Elastase | lasB | The degradation of elastin, collagen, and related matrix proteins; the spread of infection; and extracellular iron acquisition | [193] | |
| Alkaline protease | aprA | The degradation of host proteins (complement, and cytokines), the establishment of infection, and immune evasion | [194] | |
| AHL synthase | lasI | Autoinducer expression | [195] | |
| Transcriptional activator protein | Anr and Mhr | Biofilm formation under low-oxygen conditions | [84] | |
| RhlI/R | Rhamnolipids | rhlAB | The necrosis of host immune cells, biofilm formation, and immune evasion | [88,91] |
| Pyocyanin | phzABCDEFG and phzM | Oxidative stress, inflammatory response, neutrophil toxicity, the establishment of infection, and damage to host cells | [196] | |
| Hcn hydrogen cyanide (RhlR) | hcnABC | Cell toxicity and infection establishment, lung cell damage, and poor lung function | [79] | |
| Autoinducer production | lasI and rhlI | The production of AHL molecules | [197] | |
| QscR | Pyocyanin and hydrogen cyanide production | phz and hcn | Virulence factors, cell toxicity, and host cell damage | [95,96] |
| Las/Rhl quorum-sensing-dependent genes | lasIR and rhlIR and associated genes | Quorum-sensing-dependent genes | [198] | |
| TraI/R | Regulatory gene | mrtR and tmsP | Transcription regulation | [103] |
| Conjugative transfer protein | TraA, TraC, and TraD | Conjugation | [110] | |
| Type IV secretion family | trb genes | The transfer of Ti plasmid | [111] | |
| Conjugative transfer protein | trbJ and trbK | Conjugation | [112] | |
| CviI/R | Pigment production | vioA, vioB, vioC, vioD | Violacein pigment production, known for its antioxidant properties | [116,125] |
| AHL synthesis and detection | cviI cviI and cviR | AHL synthase activity to enhance the production of AHL molecules via a positive feedback loop | [116,125] | |
| Chitinase production | Chitinase genes | Degrade chitin | [119] | |
| SdiA | Cell division | ftsQ, ftsA and ftsZ | The positive regulation of the ftsQAZ gene cluster | [129] |
| Acid tolerance | GadW, and GadY | Acid tolerance in E. coli | [137] | |
| Cell attachment and biofilm formation | rck and srgE locus | Enhanced cell adhesion, invasion, and biofilm formation by enteric pathogen Salmonella enterica | [133,199] | |
| Bacterial motility | fliC and csgA | The repression of flagella and curli fimbriae | [134] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, L.; Patel, S.K.S.; Kharga, K.; Kumar, R.; Kumar, P.; Pandohee, J.; Kulshresha, S.; Harjai, K.; Chhibber, S. Molecular Mechanisms and Applications of N-Acyl Homoserine Lactone-Mediated Quorum Sensing in Bacteria. Molecules 2022, 27, 7584. https://doi.org/10.3390/molecules27217584
Kumar L, Patel SKS, Kharga K, Kumar R, Kumar P, Pandohee J, Kulshresha S, Harjai K, Chhibber S. Molecular Mechanisms and Applications of N-Acyl Homoserine Lactone-Mediated Quorum Sensing in Bacteria. Molecules. 2022; 27(21):7584. https://doi.org/10.3390/molecules27217584
Chicago/Turabian StyleKumar, Lokender, Sanjay Kumar Singh Patel, Kusum Kharga, Rajnish Kumar, Pradeep Kumar, Jessica Pandohee, Sourabh Kulshresha, Kusum Harjai, and Sanjay Chhibber. 2022. "Molecular Mechanisms and Applications of N-Acyl Homoserine Lactone-Mediated Quorum Sensing in Bacteria" Molecules 27, no. 21: 7584. https://doi.org/10.3390/molecules27217584