

Pestalotiopsis Diversity: Species, Dispositions, Secondary Metabolites, and Bioactivities

Abstract
:1. Introduction
2. Habitat and Functional Diversity of Pestalotiopsis Species
3. New Secondary Metabolites in Pestalotiopsis Species
3.1. Pestalotiopsis diploclisia


3.2. Pestalotiopsis disseminate

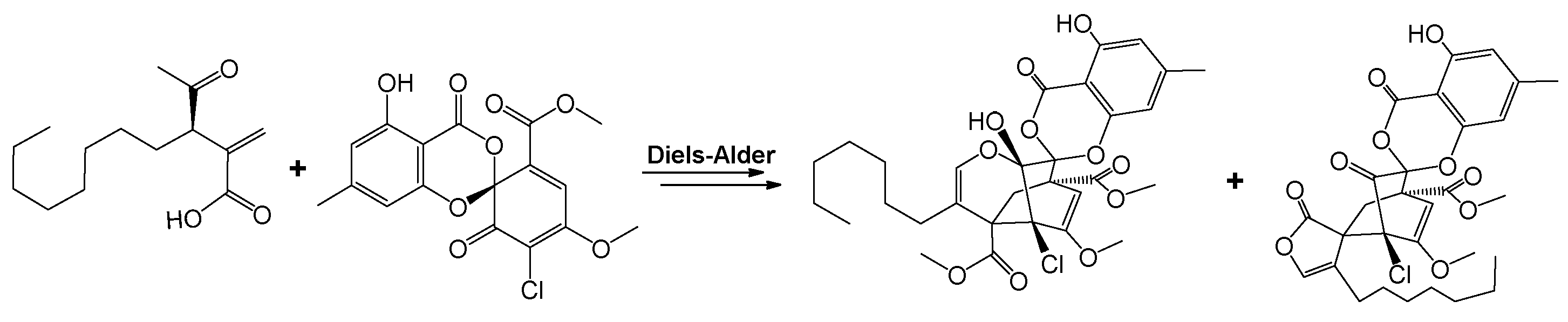
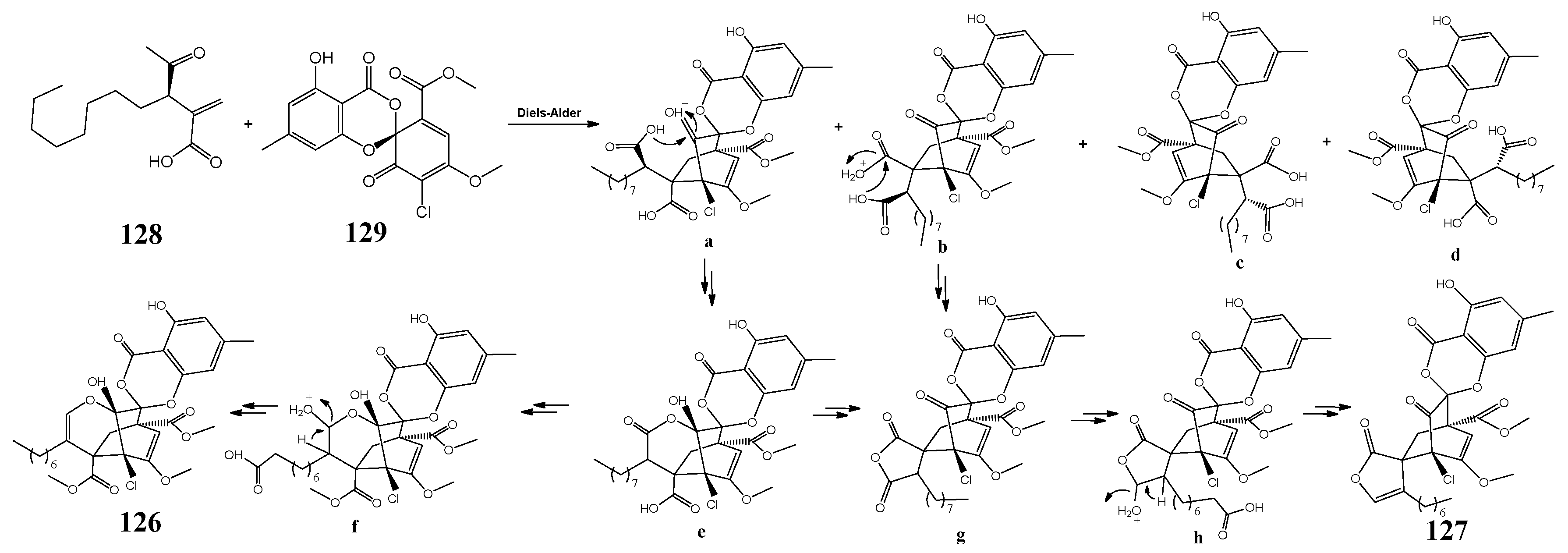
3.3. Pestalotiopsis fici

3.4. Pestalotiopsis foedan

3.5. Pestalotiopsis guepinii

3.6. Pestalotiopsis heterocornis



3.7. Pestalotiopsis humus

3.8. Pestalotiopsis karstenii

3.9. Pestalotiopsis mangiferae
3.10. Pestalotiopsis microspora





3.11. Pestalotiopsis neglecta


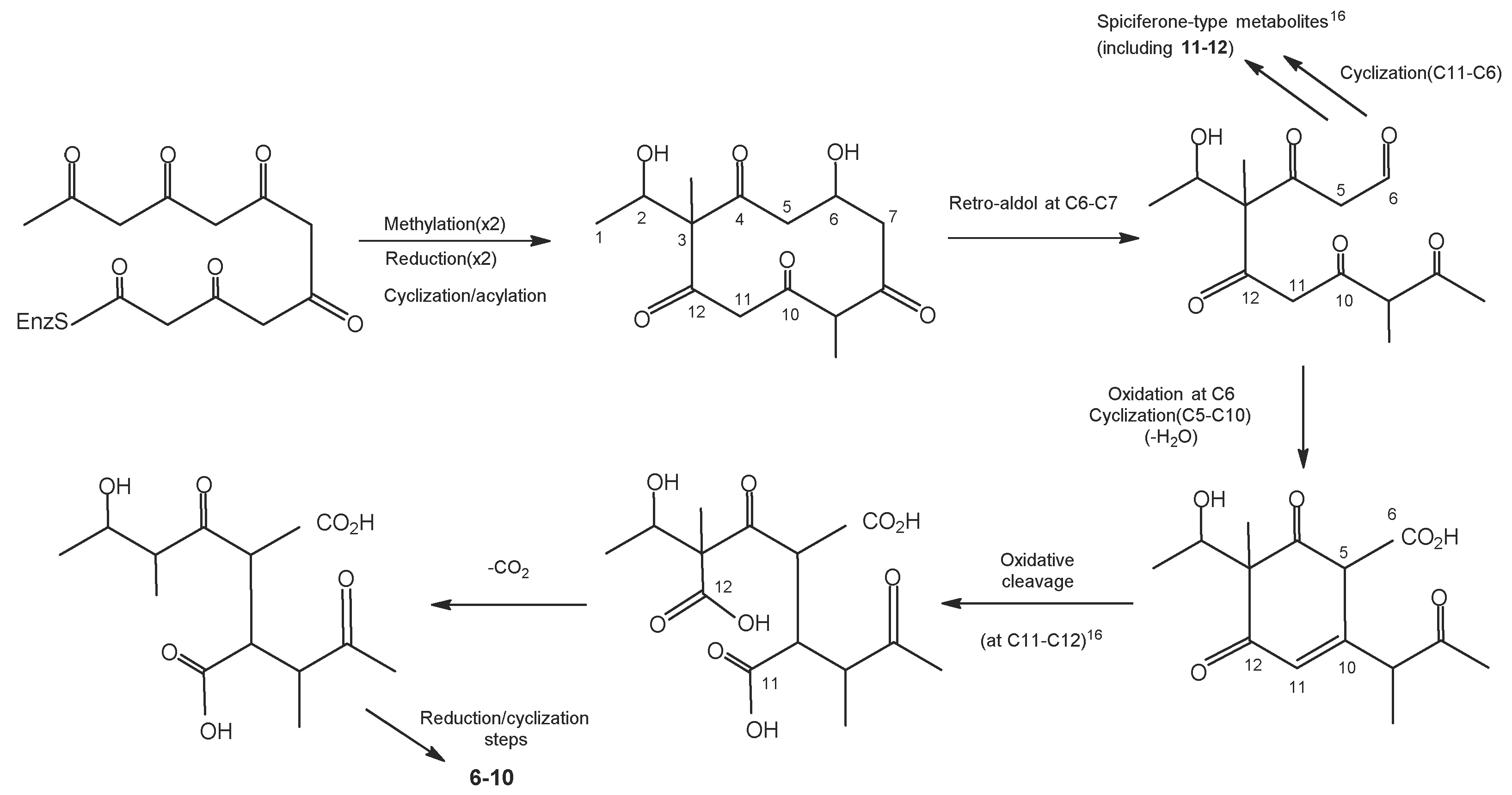
3.12. Pestalotiopsis palmarum

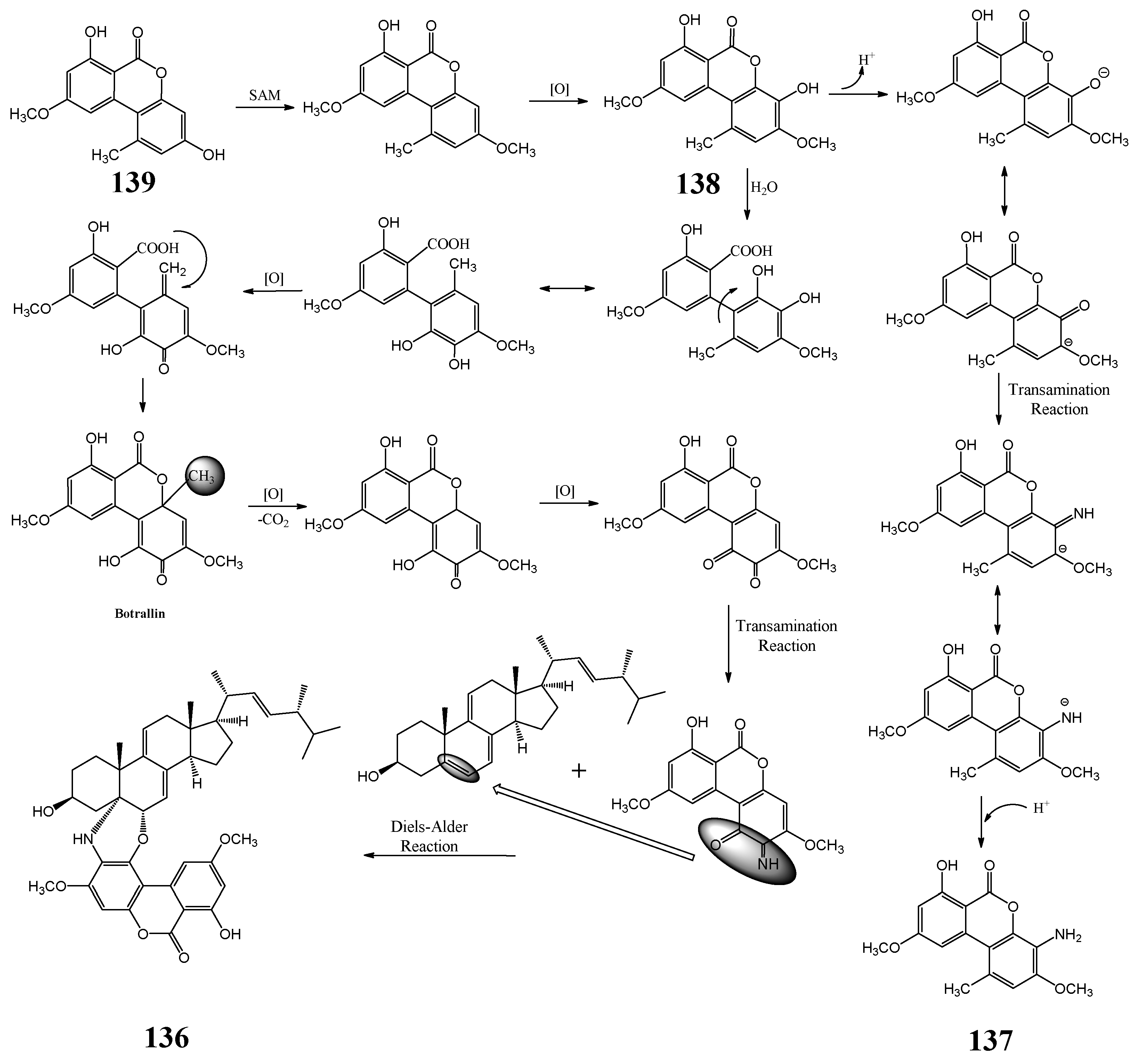
3.13. Pestalotiopsis sydowiana













4. Accurate Biosynthesis Pathways and Enhanced Accumulation of Secondary Metabolites in Pestalotiopsis
4.1. Transcription Factors Involved in Secondary Metabolite Biosynthesis in Pestalotiopsis
4.2. Histone Acetylation
4.3. Polyketide Synthases
4.4. Other Regulatory Proteins and Enzymes
5. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Subramanian, M.; Marudhamuthu, M. Hitherto unknown terpene synthase organization in taxol-producing endophytic bacteria isolated from marine macroalgae. Curr. Microbiol. 2020, 77, 918–923. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–216. [Google Scholar] [CrossRef]
- Gangadevi, V.; Muthumary, J. A Novel Endophytic taxol-producing fungus Chaetomella raphigera isolated from a medicinal plant, Terminalia arjuna. Appl. Biochem. Biotechnol. 2009, 158, 675–684. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, E.-S.R.; Zaki, A.G.; Ahmed, A.S.; Ismaiel, A.A. Production of the anticancer drug taxol by the endophytic fungus Epicoccum nigrum TXB502: Enhanced production by gamma irradiation mutagenesis and immobilization technique. Appl. Microbiol. Biotechnol. 2020, 104, 6991–7003. [Google Scholar] [CrossRef]
- Abdel-Fatah, S.S.; El-Batal, A.I.; El-Sherbiny, G.M.; Khalaf, M.A.; El-Sayed, A.S. Production, bioprocess optimization and γ-irradiation of Penicillium polonicum, as a new taxol producing endophyte from Ginko biloba. Biotechnol. Rep. 2021, 30, e00623. [Google Scholar] [CrossRef] [PubMed]
- Steyaert, R.L. Contributions à l’étude monographique de Pestalotia de Not. et Monochaetia Sacc. (Truncatella gen. nov. et Pestalotiopsis gen. nov.). Bull. Jard. Bot. Brux. 1949, 19, 285–354. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.; Guo, L.-D.; Chukeatirote, E.; Bahkali, A.; Hyde, K.D. Pestalotiopsis—Morphology, phylogeny, biochemistry and diversity. Fungal Divers. 2011, 50, 167–187. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.; Hyde, K.; Groenewald, J.; Xu, J.; Crous, P. Pestalotiopsis revisited. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Ebada, S.S.; Proksch, P. Pestalotiopsis a highly creative genus: Chemistry and bioactivity of secondary metabolites. Fungal Divers. 2010, 44, 15–31. [Google Scholar] [CrossRef]
- Xu, J.; Yang, X.; Lin, Q. Chemistry and biology of Pestalotiopsis-derived natural products. Fungal Divers. 2014, 66, 37–68. [Google Scholar] [CrossRef]
- Strobel, G.; Yang, X.; Sears, J.; Kramer, R.; Sidhu, R.S.; Hess, W.M. Taxol from Pestalotiopsis microspora, an endophytic fungus of Taxus wallachiana. Microbiology 1996, 142, 435–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, H.; Fujimoto, H.; Kimura, Y.; Hamasaki, T. Importance of the ketone function for the phytotoxicity of spiciferone A, a phytotoxin produced by the fungus Cochlioholus spicifer. Biosci. Biotechnol. Biochem. 1993, 57, 1938–1939. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Liu, H.; Niu, X.; Akhberdi, O.; Wei, D.; Wang, D.; Zhu, X. The Gα1-cAMP signaling pathway controls conidiation, development and secondary metabolism in the taxol-producing fungus Pestalotiopsis microspora. Microbiol. Res. 2017, 203, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Kathiravan, G.; Sureban, S.M. Effect of taxol from Pestalotiopsis mangiferae on A549 cells-in vitro study. J. Basic Clin. Pharm. 2010, 1, 1–9. [Google Scholar]
- Gangadevi, V.; Murugan, M.; Muthumary, J. Taxol determination from Pestalotiopsis pauciseta, a fungal endophyte of a medicinal plant. Chin. J. Biotechnol. 2008, 24, 1433–1438. [Google Scholar] [CrossRef]
- Kathiravan, G.; Raman, V.S. In vitro TAXOL production, by Pestalotiopsis breviseta—A first report. Fitoterapia 2010, 81, 557–564. [Google Scholar] [CrossRef]
- Kathiravan, G.; Sureban, S.M.; Sree, H.N.; Bhuvaneshwari, V.; Kramony, E. Isolation of anticancer drug TAXOL from Pestalotiopsis breviseta with apoptosis and B-Cell lymphoma protein docking studies. J. Basic Clin. Pharm. 2012, 4, 14–19. [Google Scholar] [CrossRef] [Green Version]
- Gangadevi, V.; Muthumary, J. Taxol production by Pestalotiopsis terminaliae, an endophytic fungus of Terminalia arjuna (arjun tree). Biotechnol. Appl. Biochem. 2009, 52, 9–15. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, Y.; Ma, X.; Wang, C.; Yue, G.; Zhang, Y.; Zhang, Y.; Li, S.; Ling, S.; Liu, X.; et al. Greater taxol yield of fungus Pestalotiopsis hainanensis from dermatitic scurf of the giant panda (Ailuropoda melanoleuca). Appl. Biochem. Biotechnol. 2014, 175, 155–165. [Google Scholar] [CrossRef]
- Li, J.-Y.; Strobel, G.; Sidhu, R.; Hess, W.M.; Ford, E.J. Endophytic taxol-producing fungi from bald cypress, Taxodium distichum. Microbiology 1996, 142, 2223–2226. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.-L.; Zhang, J.-Z.; Luo, D.-Q. The taxonomy, biology and chemistry of the fungal Pestalotiopsis genus. Nat. Prod. Rep. 2012, 29, 622–641. [Google Scholar] [CrossRef] [PubMed]
- Helaly, S.E.; Thongbai, B.; Stadler, M. Diversity of biologically active secondary metabolites from endophytic and saprotrophic fungi of the ascomycete order Xylariales. Nat. Prod. Rep. 2018, 35, 992–1014. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Stadler, M. Recent progress in biodiversity research on the Xylariales and their secondary metabolism. J. Antibiot. 2021, 74, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Sati, S.; Belwal, M. Aquatic hyphomycetes as endophytes of riparian plant roots. Mycologia 2005, 97, 45–49. [Google Scholar] [CrossRef]
- Aydoğdu, H.; Asan, A. Airborne fungi in child day care centers in Edirne City, Turkey. Environ. Monit. Assess. 2008, 147, 423–444. [Google Scholar] [CrossRef]
- Wang, J.; Liang, Z.; Li, K.; Yang, B.; Liu, Y.; Fang, W.; Tang, L.; Zhou, X. Ene-yne hydroquinones from a marine-derived strain of the fungus Pestalotiopsis neglecta with effects on liver X receptor alpha. J. Nat. Prod. 2020, 83, 1258–1264. [Google Scholar] [CrossRef]
- Wang, J.; Peng, Q.; Yao, X.; Liu, Y.; Zhou, X. New pestallic acids and diphenylketone derivatives from the marine alga-derived endophytic fungus Pestalotiopsis neglecta SCSIO41403. J. Antibiot. 2020, 73, 585–588. [Google Scholar] [CrossRef]
- Lei, H.; Zhou, X.; Hu, M.; Niu, H.; Song, C.; Chen, S.; Liu, Y.; Zhang, D. Cytotoxic polyketides from the marine sponge-derived fungus Pestalotiopsis heterocornis XWS03F09. Molecules 2019, 24, 2655. [Google Scholar] [CrossRef] [Green Version]
- Lei, H.; Lin, X.; Han, L.; Ma, J.; Dong, K.; Wang, X.; Zhong, J.; Mu, Y.; Liu, Y.; Huang, X. Polyketide derivatives from a marine-sponge-associated fungus Pestalotiopsis heterocornis. Phytochemistry 2017, 142, 51–59. [Google Scholar] [CrossRef]
- Lei, H.; Lin, X.; Han, L.; Ma, J.; Ma, Q.; Zhong, J.; Liu, Y.; Sun, T.; Wang, J.; Huang, X. New metabolites and bioactive chlorinated benzophenone derivatives produced by a marine-derived fungus Pestalotiopsis heterocornis. Mar. Drugs 2017, 15, 69. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Park, C.; Oh, E.; Sung, Y.; Lee, J.; Park, K.-H.; Kang, H. Benzophenone compounds, from a marine-derived strain of the fungus Pestalotiopsis neglecta, inhibit proliferation of pancreatic cancer cells by targeting the MEK/ERK pathway. J. Nat. Prod. 2019, 82, 3357–3365. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-F.; Liang, R.; Liao, S.-R.; Yang, B.; Tu, Z.-C.; Lin, X.-P.; Wang, B.-G.; Liu, Y. Vaccinols J–S, ten new salicyloid derivatives from the marine mangrove-derived endophytic fungus Pestalotiopsis vaccinii. Fitoterapia 2017, 120, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Parasuraman, P.; Devadatha, B.; Sarma, V.V.; Ranganathan, S.; Ampasala, D.R.; Kim, I.-W.; Patel, S.K.S.; Kalia, V.C.; Lee, J.-K.; Siddhardha, B. Inhibition of microbial quorum sensing mediated virulence factors by Pestalotiopsis sydowiana. J. Microbiol. Biotechnol. 2020, 30, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-Y.; Xue, D.-S.; Feng, X.-Y.; Yao, S.-J. Screening and production of ligninolytic enzyme by a marine-derived fungal Pestalotiopsis sp. J63. Appl. Biochem. Biotechnol. 2011, 165, 1754–1769. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, A.L.; de Medeiros, J.V.F.; Grault, C.E.; Santos, M.J.S.; Souza, A.L.A.; de Carvalho, R.W. The fungus Pestalotiopsis sp., isolated from Perna perna (Bivalvia:Mytilidae) cultured on marine farms in Southeastern Brazil and destined for human consumption. Mar. Pollut. Bull. 2020, 153, 110976. [Google Scholar] [CrossRef]
- Koh, S.; Mizuno, M.; Izuoka, Y.; Fu**o, N.; Hamada-Sato, N.; Amano, Y. Xylanase from marine filamentous fungus Pestalotiopsis sp. AN-7 was activated with diluted salt solution like brackish water. J. Appl. Glycosci. 2021, 68, 11–18. [Google Scholar] [CrossRef]
- Wikee, S.; Hatton, J.; Turbé-Doan, A.; Mathieu, Y.; Daou, M.; Lomascolo, A.; Kumar, A.; Lumyong, S.; Sciara, G.; Faulds, C.B.; et al. Characterization and dye decolorization potential of two laccases from the marine-derived fungus Pestalotiopsis sp. Int. J. Mol. Sci. 2019, 20, 1864. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.L.; Ge, H.M.; Li, F.; Song, Y.C.; Tan, R.X. New phytotoxic metabolites from Pestalotiopsis sp. HC02, a fungus residing in Chondracris rosee Gut. Chem. Biodivers. 2008, 5, 2402–2407. [Google Scholar] [CrossRef]
- Lv, C.; Huang, B.; Qiao, M.; Wei, J.; Ding, B. Entomopathogenic fungi on Hemiberlesia pitysophila. PLoS ONE 2011, 6, e23649. [Google Scholar] [CrossRef]
- Monden, Y.; Yamamoto, S.; Yamakawa, R.; Sunada, A.; Asari, S.; Makimura, K.; Inoue, Y. First case of fungal keratitis caused by Pestalotiopsis clavispora. Clin. Ophthalmol. 2013, 7, 2261–2264. [Google Scholar] [CrossRef] [Green Version]
- Magnan, R.F.; Rodrigues-Fo, E.; Daolio, C.; Ferreira, A.G.; de Souza, A.Q.L. Three highly oxygenated caryophyllene sesquiterpenes from Pestalotiopsis sp., a aungus Iiolated from bark of Pinus taeda. Zeitschrift für Naturforschung C 2003, 58, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.Q.; Zhang, L.; Shi, B.Z.; Song, X.M. Two new oxysporone derivatives from the fermentation broth of the endophytic plant fungus Pestalotiopsis karstenii isolated from stems of Camellia sasanqua. Molecules 2012, 17, 8554–8560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, H.; Wu, Y.; Ahmed, T.; Qi, X.; Li, B. Response of resistant and susceptible bayberry cultivars to infection of twig blight pathogen by histological observation and gibberellin related genes expression. Pathogens 2021, 10, 402. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-R.; ** identifies BasR, a key regulator of bacteria-triggered production of fungal secondary metabolites. eLife 2018, 7, e40969. [Google Scholar] [CrossRef]
- Brakhage, A.A.; Schroeckh, V. Fungal secondary metabolites—Strategies to activate silent gene clusters. Fungal Genet. Biol. 2011, 48, 15–22. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
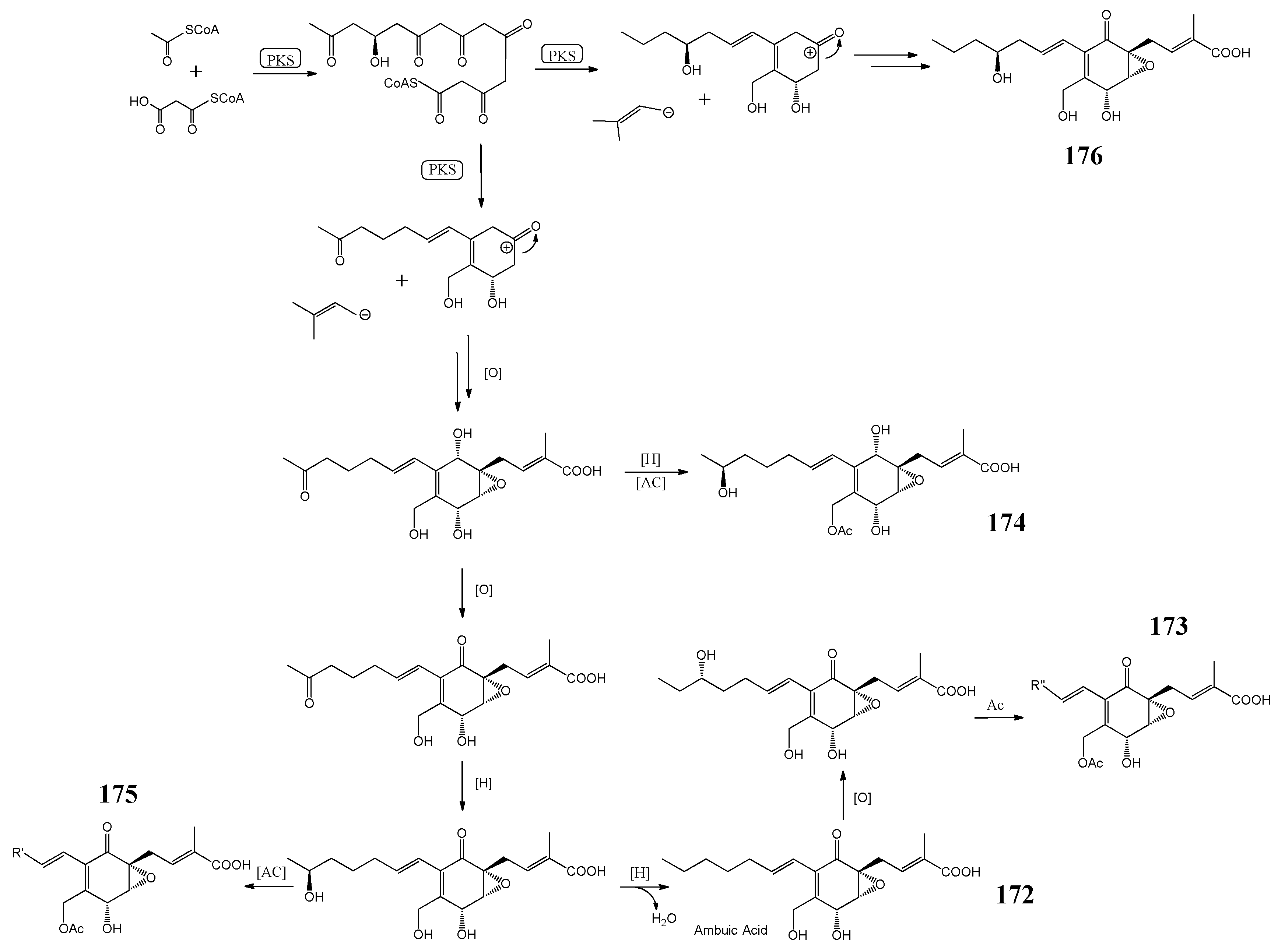{kind=link}
| Fungal Species | Metabolites | Bioactivity | References |
|---|---|---|---|
| P. adusta | pestalachlorides A–C | antifungal | [69] |
| diterpenoid | [70] | ||
| P. besseyi | furanones | [71] | |
| P. breviseta | taxol | anticancer | [16,17] |
| P. crassiuscula | a new coumarin and six known compounds | [72] | |
| P. diploclisia | pestalotioquinols G and H, pestalotioquinol A, phomonitroester, (R)-4,6,8-trihydroxy-3,4-dihydronaphthalen-1(2H)-one, and scylatone | antimalarial and cytotoxic activity | [73] |
| P. disseminata | 6-hydroxypunctaporonin E, 6-hydroxypunctaporonin B, and 6-hydroxypunctaporonin A | anti-bacteria | [74] |
| disseminins A–E, spiciferones D and E | [75] | ||
| P. fici | chloropupukeananin and chloropestolides | antimicrobial, antitumor, and anti-HIV activities | [76] |
| pestaloficiols A–E | inhibitory effects on HIV-1 replication | [77] | |
| isosulochrin, ficipyrone A, pestheic acid, iso-A82775C, pestaloficiol M, RES1214-1, and iso-A82775C | [78] | ||
| pestalofones A–E, isosulochrin, isosulochrin dehydrate, and iso-A82775C | inhibitory effects on HIV-1 replication and antifungal activity | [79] | |
| chloropestolide A | antitumor | [80] | |
| chloropupukeanolides C–E | cytotoxicity | [81] | |
| pestheic acid | [82] | ||
| chloropupukeananin | [83] | ||
| melanin | [84] | ||
| chloropupukeanolides A and B, chloropupukeanone A | anti-HIV-1 and cytotoxic activity | [85] | |
| isoprenylated chromone derivatives | [86] | ||
| Chloropestolides B–G | [87] | ||
| pestaloficins A–E | [88] | ||
| pestaloficiols Q–S | [89] | ||
| DHN melanin | [90] | ||
| 2H-pyran-2-one and 2H-furan-2-one derivatives | [91] | ||
| P. flavidula | spiroketal derivatives | cytotoxicity | [92] |
| P. foedan | pestafolide A, pestaphthalide A and B | antifungal activity | [93] |
| (−)-(4S, 8S)-foedanolide and (+)-(4R, 8R)-foedanolide | [94] | ||
| monoterpene derivatives | antifungal | [95] | |
| P. guepinii | metabolites of ciprofloxacin and norfloxacin | [96] | |
| culture broth extract | inhibit actinomycete growth | [97] | |
| pestheic acid or dihidromaldoxin | genotoxicity and mutagenicity | [98] | |
| alpha-pyrones | [99] | ||
| P. hainanensis | taxol | [19] | |
| caryophyllene-Type Sesquiterpenes | [100] | ||
| P. heterocorni | 7-hydroxy-5-methoxy-4,6-dimethyl-7-O-α-L-rhamnosyl-phthalide and 7-hydroxy-5-methoxy-4,6-dimethyl-7-O-β-D-glucopyranosyl-phthalide | [101] | |
| heterocornols A–L, methyl-(2-formyl-3-hydroxyphenyl) propanoate, cladoacetal A, xylarinol A, agropyrenol, vaccinol G, (R)-3-hydroxy-1-[(R)-4-hydroxy-1,3-dihydroisobenzofuran-1-yl]butan-2-one, and (R)-3-hydroxy-1-[(S)-4-hydroxy-1,3-dihydroisobenzo -furan-1-yl]butan-2-one | cytotoxicity and antifungal | [29] | |
| pestaloisocoumarins A and B, isopolisin B, pestalotiol A, gamahorin, pestalachloride B, pestalachloride E, pestalalactone atropisomers (8a/8b), | cytotoxicity | [30] | |
| heterocornols M and N, heterocornols O and P | anticancer | [28] | |
| P. humus | pestynol | antibacterial and antifungal activity, weak cytotoxicity | [102] |
| pestiocandin | antibacterial and antifungal activity | [103] | |
| P. jesteri | jesterone and hydroxy-jesterone | selective antimycotic activity | [104] |
| P. karstenii | pestalrone A, pestalrone B, pestalotin, hydroxypestalotin | cytotoxic activity, antiprotozoal activity | [42] |
| P. leucothës | BS, GS, and YS | Immunomodulatory | [105] |
| immunomodulatory effects | [106] | ||
| P. mangiferae | 4-(2,4,7-trioxa-bicyclo [4.1.0] heptane-3-yl) phenol | antibacterial and antifungal activity | [107] |
| P. microspora | taxol | antiproliferative activity | [11,108] |
| ɑ-pyrone | [109] | ||
| (+)-dendocarbin L, (+)-sydonic acid, and (+)-sydowic acid | cytotoxicity | [68] | |
| isopestacin | antifungal and antioxidant activities | [110] | |
| pestalotiollide B | [49,111,112,113] | ||
| pestalotiollide B, melanin | [114] | ||
| 7-epi-10-deacetyltaxol | induces apoptosis | [115] | |
| taxol, pestalotiollide B, 1, 8-dihydroxy naphthalene melanin | antitumor | [13] | |
| melanin | [116] | ||
| ursolic acid | [117] | ||
| pestalotioprolides C, D–H, 7-O-methylnigrosporolide, pestalotioprolide B, seiricuprolide, nigrosporolide, and 4,7-dihydroxy-13-tetradeca-2,5,8-trienolide | cytotoxic | [118] | |
| dibenzodioxocinons | inhibitors of cholesterol ester transfer protein | [119,120] | |
| pestalotioquinols A and B | neuroprotective | [121] | |
| pitholide E, pitholide B, pitholide D, pestalotin, PC-2, tyrosol, 4-oxo-4H-pyran-3-acetic acid | [67] | ||
| 2H-pyranone and isocoumarin derivatives | antifungal | [66] | |
| Microsporols A–C, ambuic acid | 5-lipoxygenase (5-LOX) inhibitory effects | [122] | |
| P. neglecta | ambuic acid derivatives | inhibitory activity against the NO production | [123] |
| crude methanol and ethyl acetate extract | antibacterial activity | [124] | |
| pestalotiochromenoic acids A–D, pestalotiochromones A and B | liver X receptors modulators | [125] | |
| ambuic acid | anti-inflammatory action | [126] | |
| benzophenones | inhibit pancreatic cancer cells | [31] | |
| Ene-yne Hydroquinones | [26] | ||
| pestalopyrones A–D | [127] | ||
| pestallic acids F and G, pestalotiopyrone N, neopestalone, sesquicaranoic acid B, monocycloalternarene B, pestalone, 2,4-dihydroxy-3,5,6-trimethyl benzoic acid, and citreorosein | [27] | ||
| P. palmarump | sinopestalotiollides A–D, 3′-O-methyldehydroisopenicillide, ∆1′3′-1′-dehydroxypenicillide, dehydroisopenicillide, 2′-hydroxy-3′,4′-didehydropenicillide, scirpyrones A, 5,6-dihydro-4-methoxy-2H-pyran-2-one, LL-P880α, (6S,1′S,2′R)-LL-P880b, photipyrone B, PC-2, (1′S,2′R)-LL-P880γ, necpyrone C | cytotoxic | [128] |
| P. pauciseta | taxol | anticancer | [15] |
| P. photiniae | phthalide derivatives | against plant pathogens | [129] |
| photipyrones A, B, C | modest inhibitory effects on the growth of MDA-MB-231 | [130] | |
| 4-(3′,3′-dimethylallyloxy)-5-methyl-6-methoxyphthalide | induced G1 cell cycle arrest and apoptosis in a dose-dependent manner | [131] | |
| 4-(3′,3′-Dimethylallyloxy)-5-methyl-6-methoxy-phthalide | cytotoxicity | [132] | |
| three new phthalide derivatives and six known phthalide derivatives | antifungal activities | [133] | |
| photinides A–F | cytotoxic | [52] | |
| Pestalotiopsis sp. | RES-1214-1 and -2 | non-peptidic endothelin type A receptor antagonists | [134] |
| Pestalotiopsis sp. | pestalotiopamide E | [135] | |
| Pestalotiopsis sp. | pestalotiopens A and B | [136] | |
| Pestalotiopsis sp. | pestalactams A–C | [137] | |
| Pestalotiopsis sp. | pestaloficiol J, (±)-pestaloficiol X | [138] | |
| Pestalotiopsis sp. | pestaloquinols A and B | [139] | |
| Pestalotiopsis sp. | demethylincisterol A3, dankasterone B, (22E, 24R)-ergosta-7,9, 22-triene-3β, 5α, 6α-triol, ergosta-5,7,22-trien-3-ol, 5,8-epidioxy-5,8-ergosta-6,22E-dien-3-ol, stigmastan-3-one, stigmast-4-en-3-one, stigmast-4-en-6-ol-3-one, flufuran, (2-cis, 4-trans)-abscisic acid, similanpyrone B. | induce G0/G1 cell cycle arrest and apoptosis in human cancer cells | [140] |
| Pestalotiopsis sp. | pestaloporonins | [141] | |
| Pestalotiopsis sp. | polyketide-terpene hybrid metabolites | [142] | |
| Pestalotiopsis sp. | pestaloporinates A–G and 14-acetylhumulane | [143] | |
| Pestalotiopsis sp. | isocoumarin derivatives | antifungal | [144] |
| Pestalotiopsis sp. | pestalotiopsolide A, taedolidol and 6-epitaedolidol | [41] | |
| Pestalotiopsis sp. | (±)-pestalachloride D | antibacterial | [145] |
| Pestalotiopsis sp. | cytosporones J-N, pestalasins A-E, pestalotiopsoid A, cyclosporine C, dothiorelone B, and 3-hydroxymethyl-6,8-dimethoxycoumarin (13). | [146] | |
| Pestalotiopsis sp. | (+)- and (−)-pestaloxazine A | antiviral | [147] |
| Pestalotiopsis sp. | pestalotiopsins A and B | immunosuppressive | [148] |
| Pestalotiopsis sp. | ambuic acid and torreyanic acid derivatives | antimicrobial activity | [149] |
| Pestalotiopsis sp. | pestalotiopsones A–F | moderate cytotoxicity | [150] |
| Pestalotiopsis sp. AcBC2 | pestalols A–E, 4-hydroxyphenethyl 2-(4-hydroxyphenyl) acetate, r-hydroxyphenyl acetic acid methyl ester, transharzialactones A and F, 3-hydroxy-3-methyl-d-lactone, 3β,5α, 9α-trihydroxy-7, 22-en-ergost-6-one, and 3b-hydroxy-sterol | cytotoxicity, inhibitory activities against Influenza A virus subtype (H3N2), Swine Flu (H1N1) viruses, tuberculosis | [151] |
| Pestalotiopsis sp. BC55 | exopolysaccharide | [152] | |
| Pestalotiopsis sp. cr013 | pestalpolyols A–D | cytotoxic | [153] |
| Pestalotiopsis sp. cr014 | pestalotic acids A–I | antibacterial | [154] |
| Pestalotiopsis sp. PG52 | pestalpolyols E–H | cytotoxic activities | [155] |
| Pestalotiopsis sp. EJC07 | (4S)-4,8-dihydroxy-1-tetralone, uracil, uridine, p-hydroxybenzoic acid, ergosterol, ergosterol peroxide, cerevisterol and ducitol | [74] | |
| Pestalotiopsis sp. FT172 | pestallic acids A–E | anti-proliferative | [156] |
| Pestalotiopsis sp. HC02 | pestalotines A and B | [38] | |
| Pestalotiopsis sp. HHL101 | pestalotiopisorin B | [157] | |
| Pestalotiopsis sp. HQD-6 | pestalotiopin B and pestalotiopyrone N | very weak cytotoxic | [158] |
| Pestalotiopsis sp. IQ-011 | cuautepestalorin, cytosporin M, cytosporin N, oxopestalochromane, pestalone | inhibitory properties against α-glucosidase from S. cerevisiae | [159] |
| Pestalotiopsis sp. M-23 | drimane sesquiterpenoids, 2α-hydroxyisodrimeninol, a new isochromone derivative | weak antibacterial | [160] |
| Pestalotiopsis sp. PSU-MA69 | pestalochromones A–C, pestalotethers A–D, pestaloxanthone, pestalolide | antifungal activity against Candida albicans and Cryptococcus neoformans | [161] |
| Pestalotiopsis sp. Z233 | 1β,5α,6α,14-tetraacetoxy-9α-benzoyloxy-7β H-eudesman-2β,11-diol and 4α,5α-diacetoxy-9α-benzoyloxy-7βH-eudesman-1β,2β,11,14-tetraol | tyrosinase inhibitory activities | [162] |
| Pestalotiopsis spp. | ambuic acid | antifungal | [163] |
| Pestalotiopsis spp. | chromones, cytosporones, polyketides, terpenoids and coumarins | [164] | |
| P. sydowiana | 1-O-methyldehydroisopenicillide, pestalotiollide B, pestalotiollide A, dehydroisopenicillide, 6-hydroxymethyl-4-methoxy-5,6-dihydro-2H-pyran-2-one, pestalotiopyrone D, pestalotiopyrone E, pestalotiopyrone G, LL-P880b, and photipyrone B | 20S proteasome inhibitory activities | [165] |
| cyclo(-Leu-Pro) and 4-hydroxyphenylacetamide | antimicrobial, | [33] | |
| P. terminaliae | taxol | anticancer | [18] |
| P. theae | chloroisosulochrin, ficipyrone A and pestheic acid | strong activity against respiratory syncytial virus | [166] |
| pesthetoxin | [167] | ||
| pestalotheols A–D | inhibitory effect on HIV-1 LAI replication | [168] | |
| pestalazines and pestalamides | inhibitory effects on HIV-1 replication and antifungal activity | [169] | |
| chlorotheolides A and B, 1-undecene-2,3-dicarboxylic acid, maldoxin | [170] | ||
| pestalotiones A–D | [171] | ||
| chloropupukeananin and pestalofone C | regulate autophagy through AMPK and Glycolytic Pathway | [172] | |
| pestathenols A and B, pestatheranone A, punctaporonins O, P, and R, ficifuranone B, and decarestrictine D | [173] | ||
| pestalothenin A–C | cytotoxic and antibacterial activity | [174] | |
| P. uvicola | pestauvicomorpholine A and three alternariol analogues | cytotoxicity against mouse melanoma (B16-BL6) cell line | [175] |
| bilobalide | suppresses adipogenesis in 3T3-L1 adipocytes via the AMPK signaling pathway; protect BV2 microglia cells against OGD/reoxygenation injury | [176,177,178] | |
| P. vaccinii | vaccinols J-S, trans-sordarial, trans-sordariol, cis-sordariol, 4-hydroxyphthalide, pestalotiopin A | [32] | |
| P. versicolor | 4,6-dihydroxy-7-formyl-3-methyl coumarin, 6-[(7S,8R)-8-propyloxiran-1-yl]-4-methoxy-pyran-2-one | devoid of significant antifungal activity against F. solani, Ustilago maydis, and C. albicans | [179] |
| P. virgatula | pestalospiranes A and B | [180] | |
| 9-hydroxybenzo[C]oxepin-3[1H]-one | [181] | ||
| virgatolides A–C | [182] | ||
| P. yunnanensis | pestalotic acids A–G | significant antimicrobial activity | [183] |
| P. zonata | pestalrones A and B, pestalotin, hydroxypestalotin, pestazonatic acid, necpyrones A–B | [184] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.; Wang, Y.; Yang, Y. Pestalotiopsis Diversity: Species, Dispositions, Secondary Metabolites, and Bioactivities. Molecules 2022, 27, 8088. https://doi.org/10.3390/molecules27228088
Wu C, Wang Y, Yang Y. Pestalotiopsis Diversity: Species, Dispositions, Secondary Metabolites, and Bioactivities. Molecules. 2022; 27(22):8088. https://doi.org/10.3390/molecules27228088
Chicago/Turabian StyleWu, Chu, Yun Wang, and Yujie Yang. 2022. "Pestalotiopsis Diversity: Species, Dispositions, Secondary Metabolites, and Bioactivities" Molecules 27, no. 22: 8088. https://doi.org/10.3390/molecules27228088