1. Introduction
High-altitude hypoxia exerts a great impact on the physical and mental health of plateau residents. It is known that when the human body rises to a plateau area above 3000 m, the partial pressure of oxygen in the atmosphere will drop sharply with the increase in altitude, which results in hypoxia of tissue cells. As part of these changes, oxygen demand often exceeds supply and can result in overall oxygen deficits [
1,
2,
3,
4,
5]. This condition indeed seriously affects the body’s metabolism and normal physiological functions, including acute altitude sicknesses such as high altitude pulmonary edema (HAPE) and high altitude cerebral edema (HACE) [
1,
6,
7]. This stressful condition could also exacerbate risk of cystemia and various chronic cardiovascular diseases [
2]. The changes in the body under high altitude hypoxia exposure and the molecular mechanisms of various tissue systems in response to hypoxia stress have become the subject of high-altitude medical research in recent years.
The immune system is an important defense system of the body. The immune system plays an irreplaceable role in maintaining the homeostasis of tissues. Accumulative studies reported that hypoxia regulates the proliferation, development and effector function of immune cells through transcriptional regulation driven by hypoxia inducible factors (HIFs) [
3,
4]. It has been reported that hypoxia could increase the immune cell infiltration and inflammatory response in various organs of mice, and hence leads to the occurrence of acute altitude sickness [
5,
8]. Therefore, the relative impact of high-altitude hypoxic environment on the body immune system urgently requires widespread attention.
Arachidonic acid (AA) is an important polyunsaturated fatty acid, which represents a wide range of immune-physiological effects in animals. As an extremely important structural lipid, it widely exists in mammalian organs, muscles and blood. For example, in tissues, it is an essential fatty acid in animals. Arachidonic acid is the precursor of various biologically active substances [
9]. There are mainly three types of enzymes involved in the metabolism of arachidonic acid. The oxidation reactions that catalyze the metabolism of arachidonic acid mainly include: epoxidation, allyl oxidation and Ω hydroxylation [
10]. AA is catalyzed by lipoxygenase (LPO) to generate hydroxyeicosatetraenoic acid (HETEs), leukotrienes (LT) and lipoxins (LXs) [
11], and LPO is a dioxygenase [
12]. The LPO metabolites of arachidonic acid (HETEs and their precursors HPETEs and LTs) have inhibitory effects on immune cells and immune responses. In addition, the synthesis of arachidonic acid as a substrate under the catalysis of enzymes has biological activity. In this regard, given the arachidonic acid substrate is reduced or replaced, this will directly lead to the change in some hypoxia induced regulators, resulting in a series of inflammatory and immune cell responses [
13]. However, the metabolic pathways of arachidonic acid are complex, and it is difficult to identify the key factors that affect the immune response, and the physiological function of arachidonic acid in different stressful conditions remain unclear [
11].
In recent years, multi-omics combined analysis technology has been gradually applied to the study of key factor analysis of complex biological pathways. Various effective protein factors, biomarkers and anticancer factors were successfully identified through combined analysis of transcriptome, proteome and metabolome. For example, based on proteomic and transcriptomic integrative analysis, Wang and colleagues reported that total m6A levels of mRNAs were reduced during hypoxia, possibly mediated by induction of the m6A eraser ALKBH5, and they suggested that crosstalk between m6A and HIF1 pathways is important for cells to be deficient in hypoxia [
14]. In addition, integrative analysis of proteomics and metaboliomics provides the potential to identify changes in protein and metabolites levels following hypoxic exposure and may provide important insights into the mechanisms involved in hypoxic preconditioning [
15,
16]. The reaction of oxygen is crucial. Through combined proteomic and metabolomic analysis, some scholars have reported the effect of KDAC inhibition (KDACi) on the metabolic profile, and pointed out that the up-regulation of glycolysis, TCA cycle, oxidative phosphorylation and fatty acid synthesis has become a common metabolic response of KDACi [
17]. Through combined transcriptome and metabolome analysis, it was found that amino acids not only act as potential osmotic agents, but also contribute to energy generation in response to hypoxia in the clam hypoxia response [
18].
In this study, we focused on identifying which key molecular factors are involved in immunological responses in the hypoxia response. With this purpose, we applied a combined proteomic and metabolomic analysis to identify co-altered metabolic and biological pathways. The key differential proteins and/or metabolites in the metabolic pathway of arachidonic acid will be obtained to describe the molecular mechanism that affects the immune response of kidney organs, which is beneficial to expand our knowledge on main physiological functions of arachidonic acid in different organs, and bring new insights for the spleen targeted rational design upon arachidonic acid metabolism of new therapies.
2. Results
In this study, we first performed a TMT-based proteomics analysis, and results show that there are 72,824 peptides detected and these peptides were mapped to 7727 proteins (
Table S2). The peptides numbers declined gradually following the protein coverage from 0.1~1.0 (
Figure S1A). The cumulative fractions increased dramatically when coefficient of variation were between 0.1 to 0.4, and appear to be plateau when CV was more than 0.4 for either hypoxia group or normalxia group (
Figure S1B). According to subcellular localization analysis, the 32.97% proteins were annotated to be related to nuclear proteins accounting for total 7727 proteins (
Figure S1C). The length values for the most peptides were ranged from 8 to 16 (
Figure S2A). The protein mass ranges majority at from 30~60 kDa (
Figure S2B). The most mass spectrum shows desirable precursor ion tolerance (
Figure S2C). The ratio of unique peptides increased gradually when the peptide numbers were less than 28 (
Figure S2D). This evidence suggests that the proteomics are successfully performed in this study.
According to GO and KEGG analysis, we found that most proteins are related to oxidation–reduction process and protein phosphorylation. Around 227 and 197 proteins are related to nucleus and integral component of membrane. Most proteins are related to protein binding and ATP binding based on molecular function classification (
Figure S3A). In addition, the proteins annotated were involved in many primary metabolic processes or their relevant derivate metabolic pathways, such as carbon hydrate metabolism, immune system, environmental adaptation, transport catabolism and signal transduction (
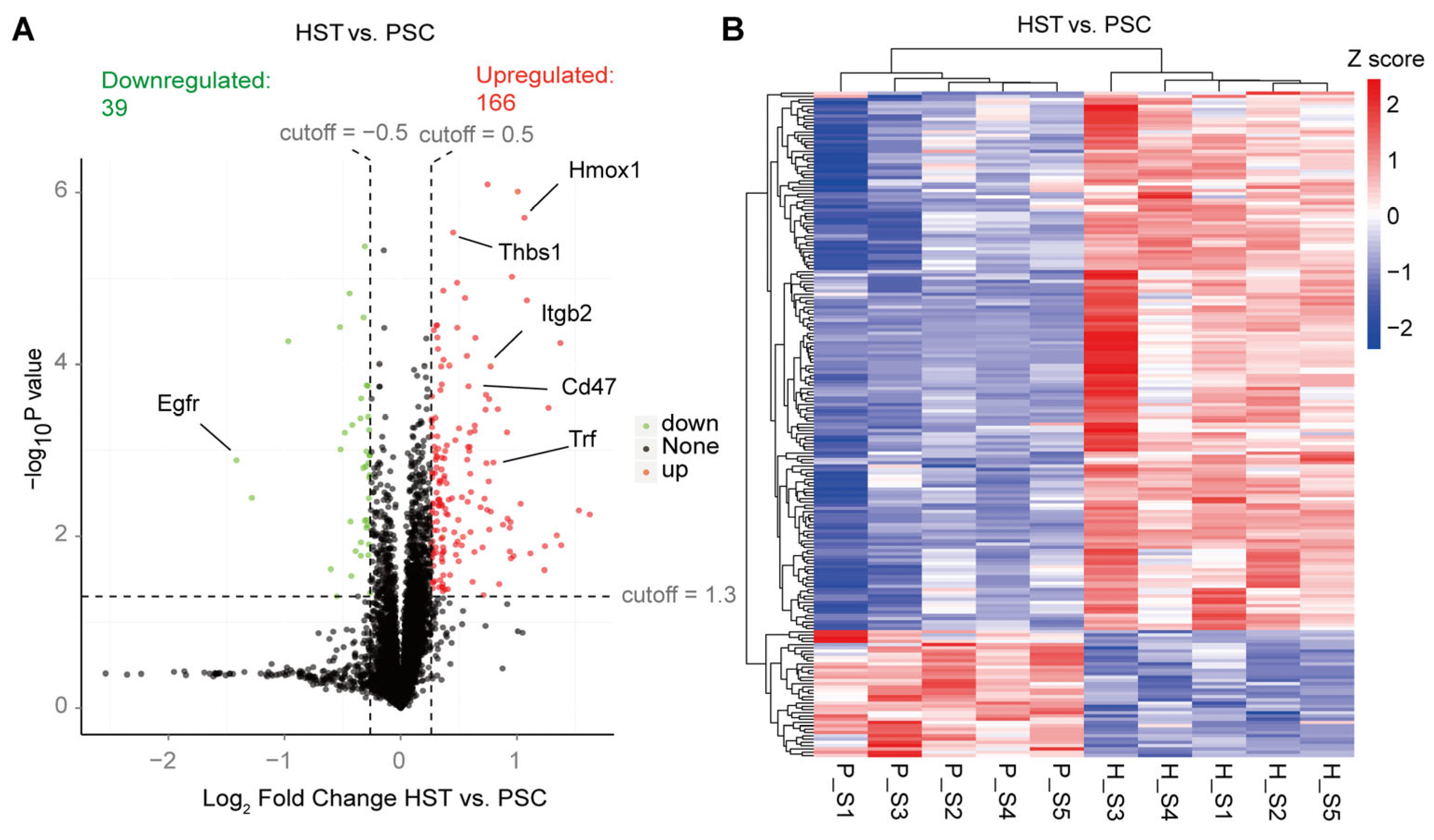
Figure S3B). In addition, we found that there are 166 upregulated differentially expressed proteins (DEPs) induced by hypoxia treatment in the spleen of mice, which is four times more than the numbers of downregulated DEPs (
Figure 1A;
Table S2). These DEPs contain some known hypoxia markers that are upregulated in the hypoxia treatment mice, including Hmox1, Trf, Thbs1, Itgb2 and Cd47 (
Figure 1A;
Table S3). All DEPs were visualized based on the heatmap indicated in
Figure 1B.
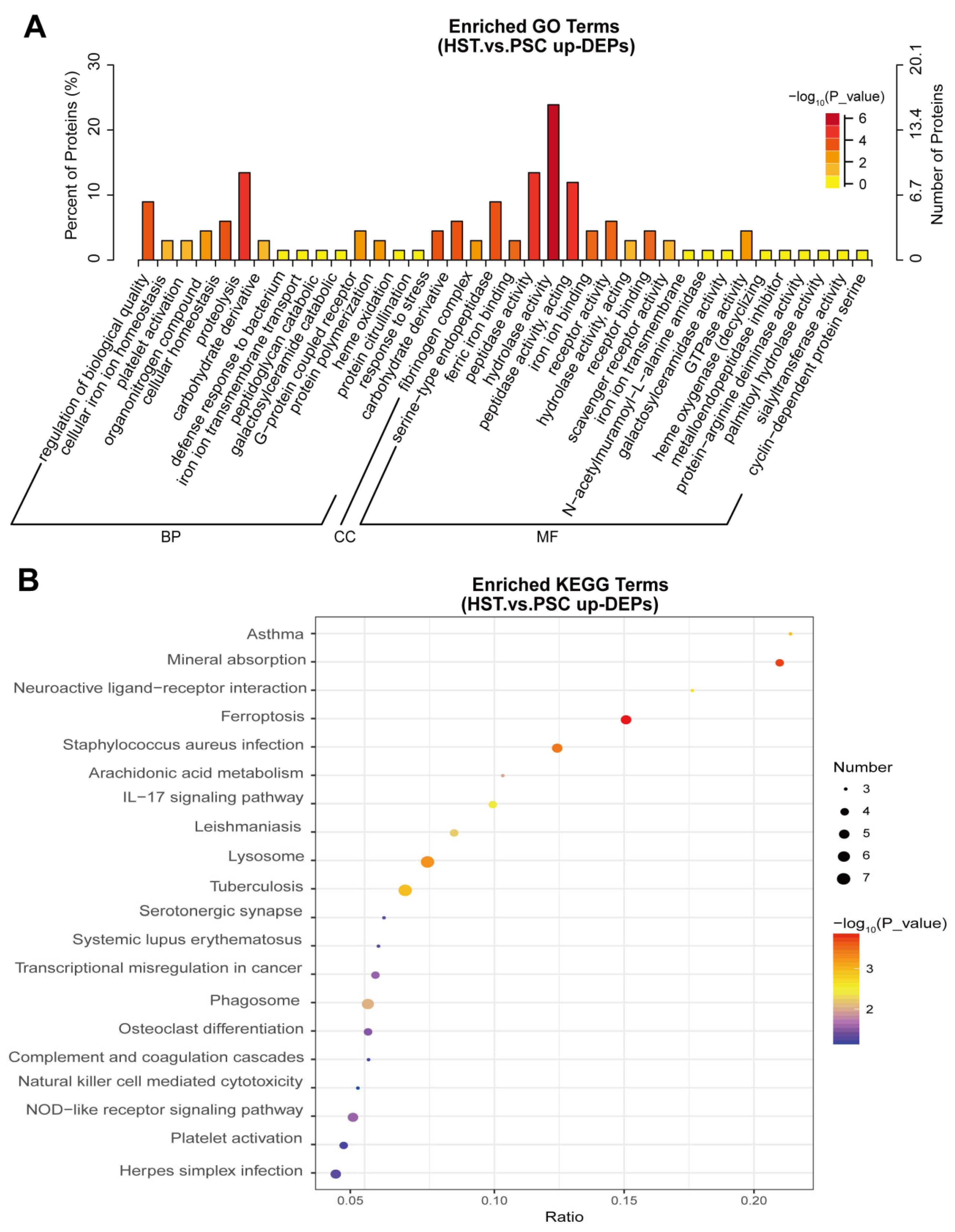
Among the 166 upregulated DEPs, we further performed the GO and KEGG analysis to found the altered biological pathways and metabolic pathways. Results show that proteolysis, carbohydrate derivative catabolic process, hydrolase activity and GTPase activity were significantly enriched in the list of these proteins (
Figure 2A). In addition, we found that mineral absorption, neuroactive ligand–receptor interaction, arachidonic acid metabolism, IL-17 signaling pathway and NOD-like preceptor signaling pathway were significantly enriched in the list of 166 upregulated DEPs (
Figure 2B).
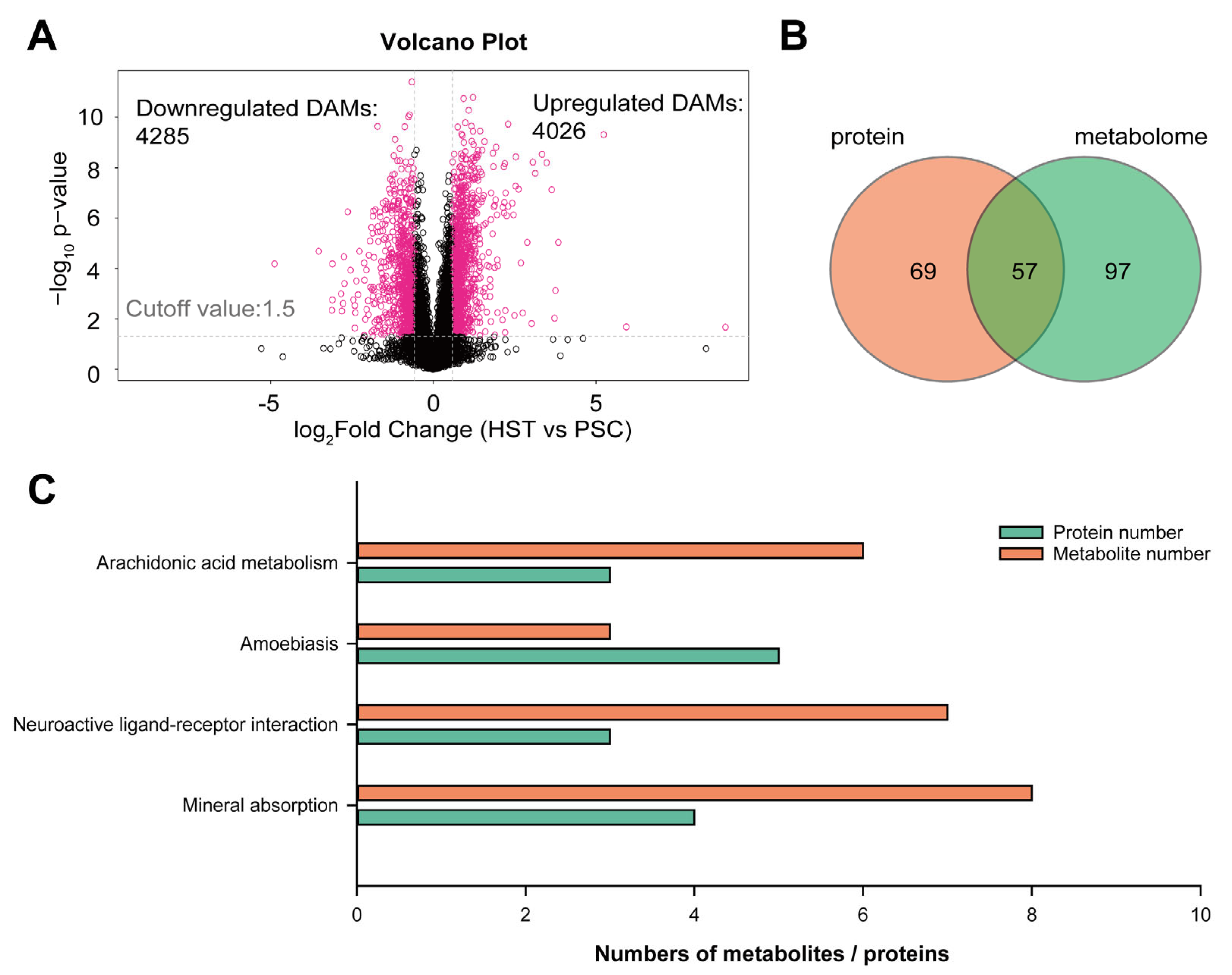
Therefore, we are interested to carry out an integrative analysis of proteomic and non-targeted metabolism to found if there are some overlapped metabolic pathways that co-occurred in both analyses. From the metabolomic analysis, we identified 4285 downregulated differentially abundant metabolites (DAMs) and 4026 upregulated DAMs (
Figure 3A), while there are 74 and 76 downregulated DAMs and upregulated DAMs with annotation by metabolic library (
Table S4). Next, we attempted to compare the overlapped metabolic pathways that significantly enriched in the list of DEPs and DAMs. Result show that there are 57 overlapped metabolic pathways that co-occurred in the list of DEPs and DAMs (
Figure 3B;
Figure S4;
Table S5). The top four KEGG terms that were both significantly enriched in the list of DEPs and DAMs include arachidonic acid metabolism, amobiasis, neuroactive ligand–receptor interaction and mineral absorption (
Figure 3C).
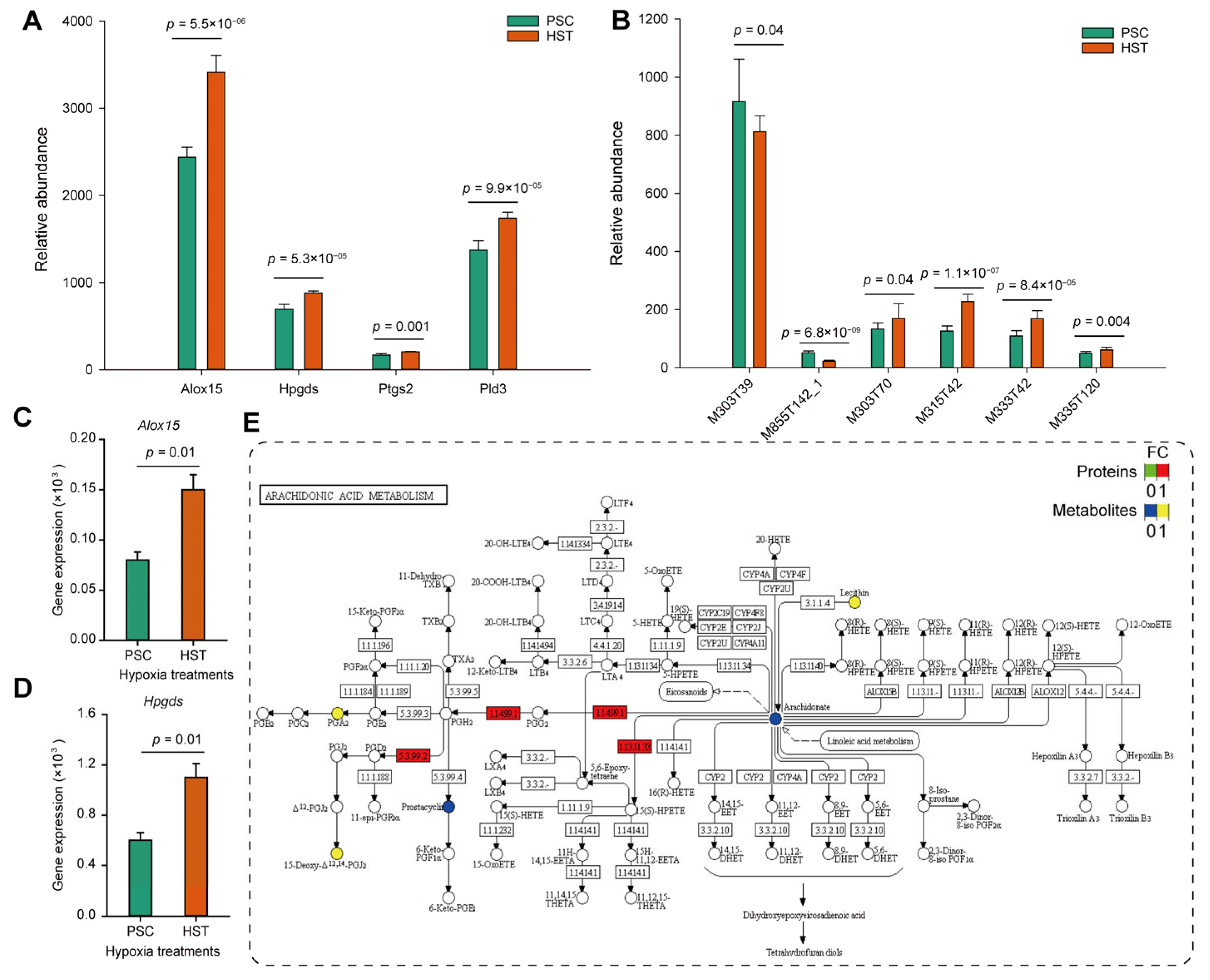
Furthermore, we compared the expression levels of proteins and metabolites involved in arachidonate acid metabolism, and found that arachidonate 15-lipoxygenase (Alox15), hematopoietic prostaglandin D synthase (Hpgds), prostaglandin G/H synthase (Ptgs2) and phospholipase D (Pld3) were significantly upregulated regarding their protein abundance from proteomics analysis (
Figure 4A). Except for the arachidonic acid (peroxide free) (M303T39) and 1-stearoyl-2-oleoyl-sn-glycerol 3-phosphocholine (M855T142_1), 20-hydroxyarachidonic acid (M303T70), 15-deoxy-delta-12,14-PGJ2 (M315T42), 8-iso-prostaglandin A2 (M333T42) and prostaglandin D2 (M335T120) were all up-regulated by hypoxia treatments (
p < 0.05) (
Figure 4B). Then the qPCR experiments were conducted to determine the mRNA expression levels. Results show that the expression levels of
Alox15 and
Hpgds were upregulated by one and four times resulted by hypoxia treatments, respectively (
Figure 4C,D). The expression levels of
Ptgs2 gene were slightly enhanced but not significantly altered due to hypoxia effects (
Figure S5). Consistently, the protein expression of Hpgds, Alox15 and Ptgs2 were increased by at least two times based on Western blot experiments (
Figure S6).
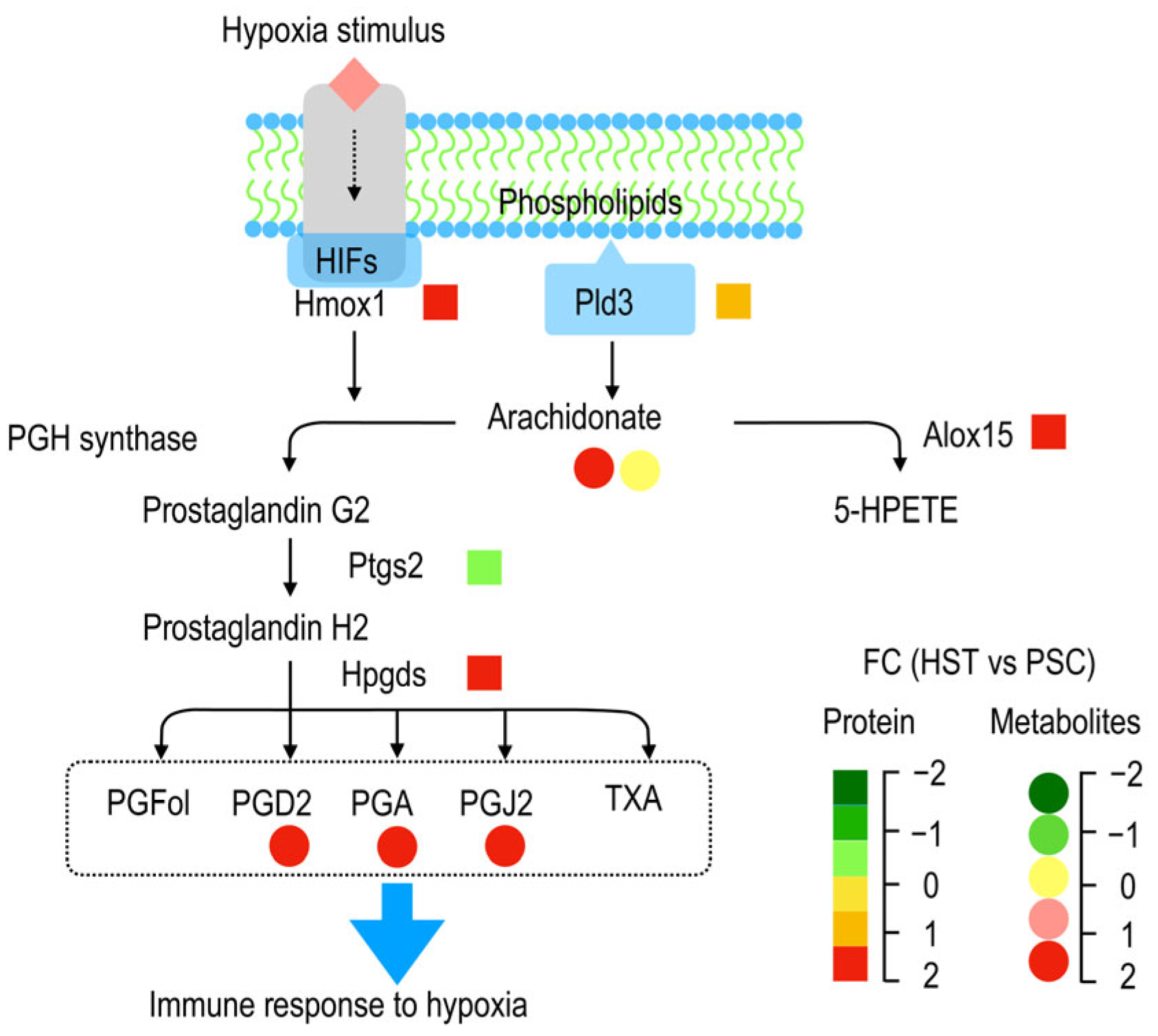
As summarized by
Figure 5, membrane phospholipids were converted to arachidonate acid catalyzed by Pld3, and the two forms of latter metabolite (arachidonate acid), i.e., arachidonate acid (peroxide free, M303T39) and 20-hydroxyarachidonic acid (M303T70), as detected in current study were not altered or significantly enriched by hypoxia treatments, and they can be converted to both prostaglandin G2 and 5-HPETE by PGH synthase and Alox15, respectively. Hypoxia stimulates expression of some genes related to hypoxia induced factors (HIFs) such as Hmox1 (heme oxygenase 1, ENSMUSP00000005548.6), which could participate in the prostaglandin G2 biosynthesis (
Table S3). The prostaglandin G2 was than sequentially converted to prostaglandin H2 and other derivatives such as PGFol, PGD2, PGA, PGJ2 and TXA. PDG2 (M335T120), PGA (M333T42) and PGJ2 (M315T42) as detected by metabolomic analysis show significantly enriched, and these metabolites are hypothetically helpful to promote immune response to hypoxia stress.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




