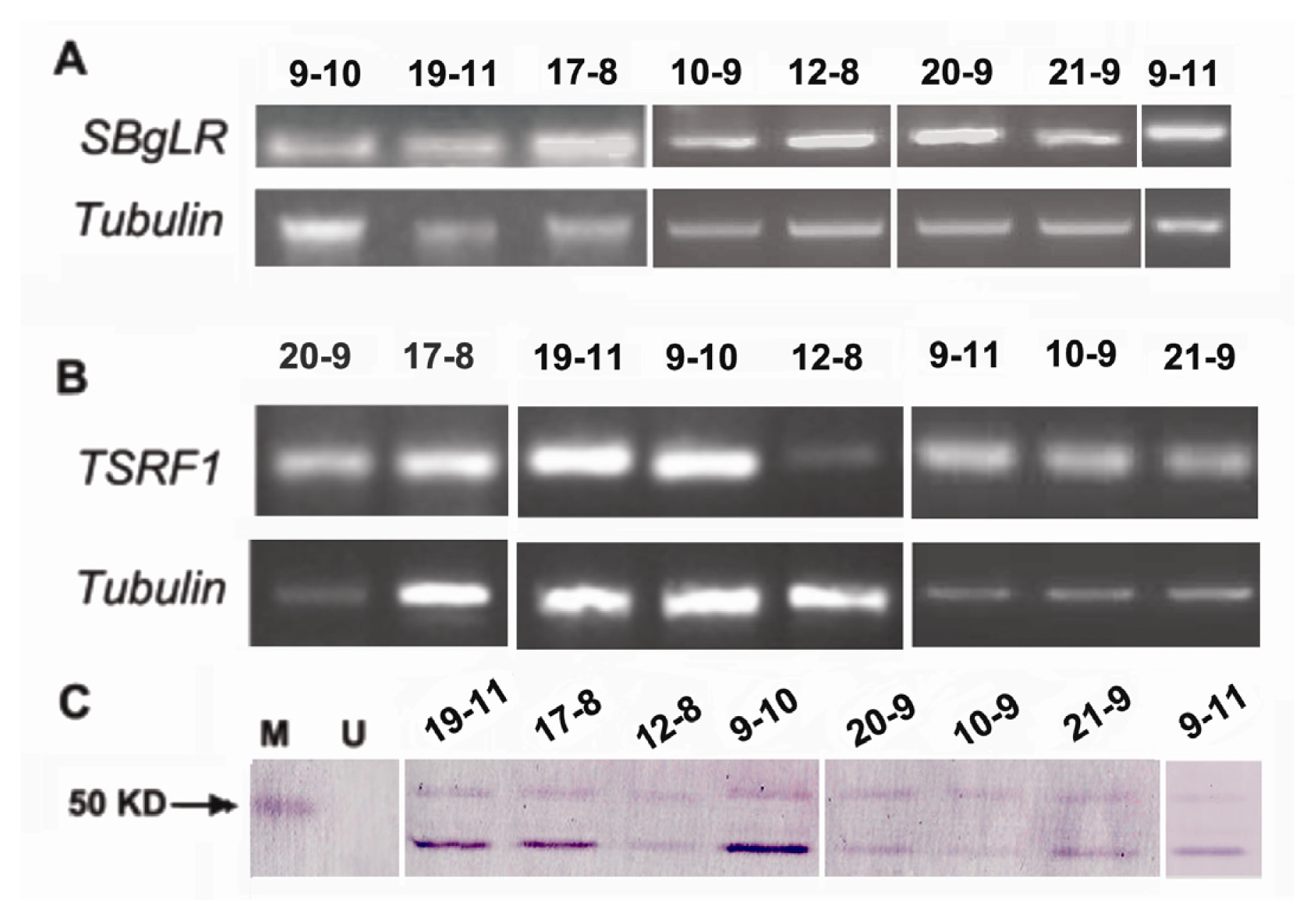
The overexpression of
TSRF1 increases pathogen tolerance in tobacco and tomato [
29] and drought resistance in rice [
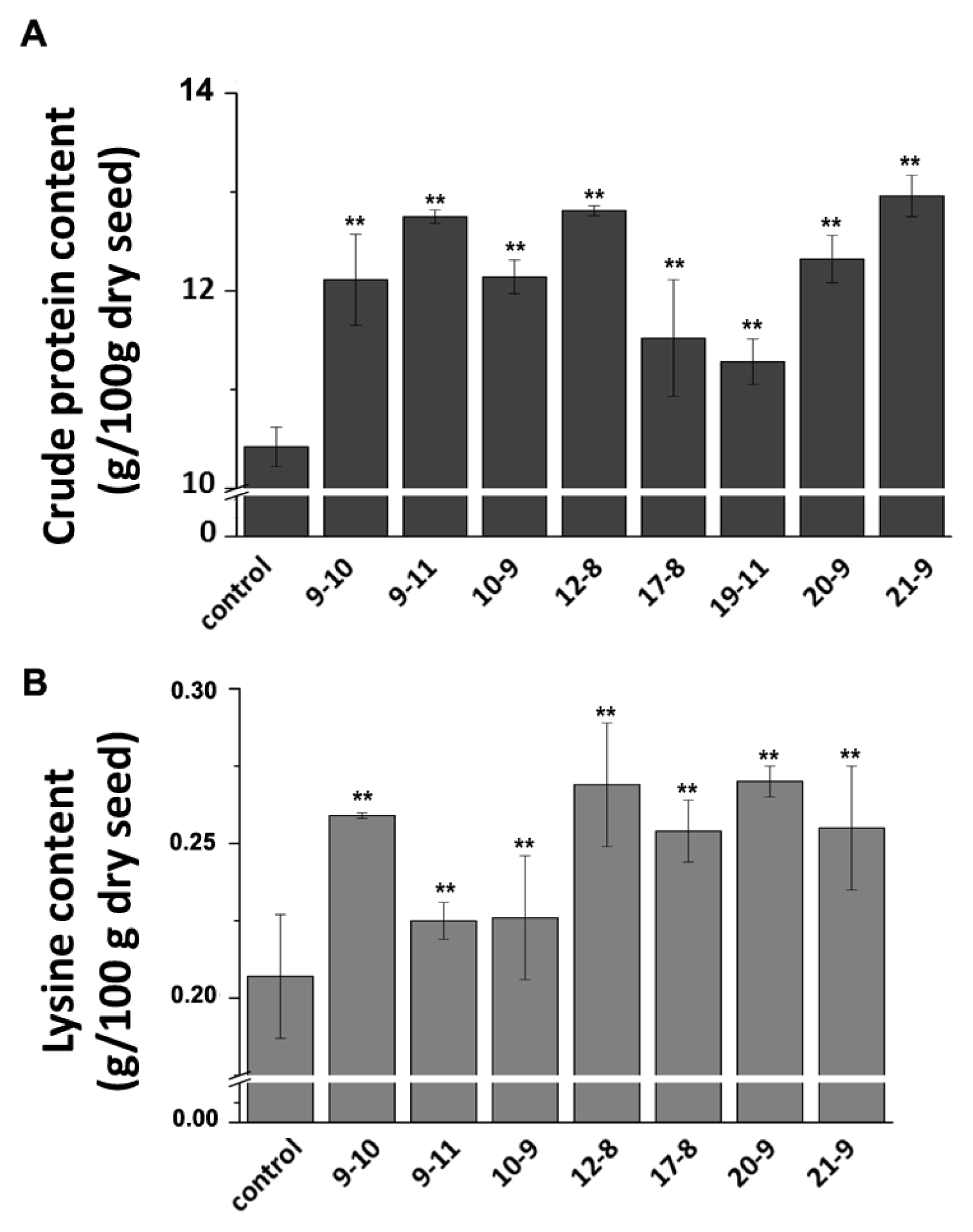
32]. To investigate whether TSRF1 have salt tolerance in transgenic maize, four-leaf stage seedlings of three transgenic lines were used for salt treatment. The result was shown in
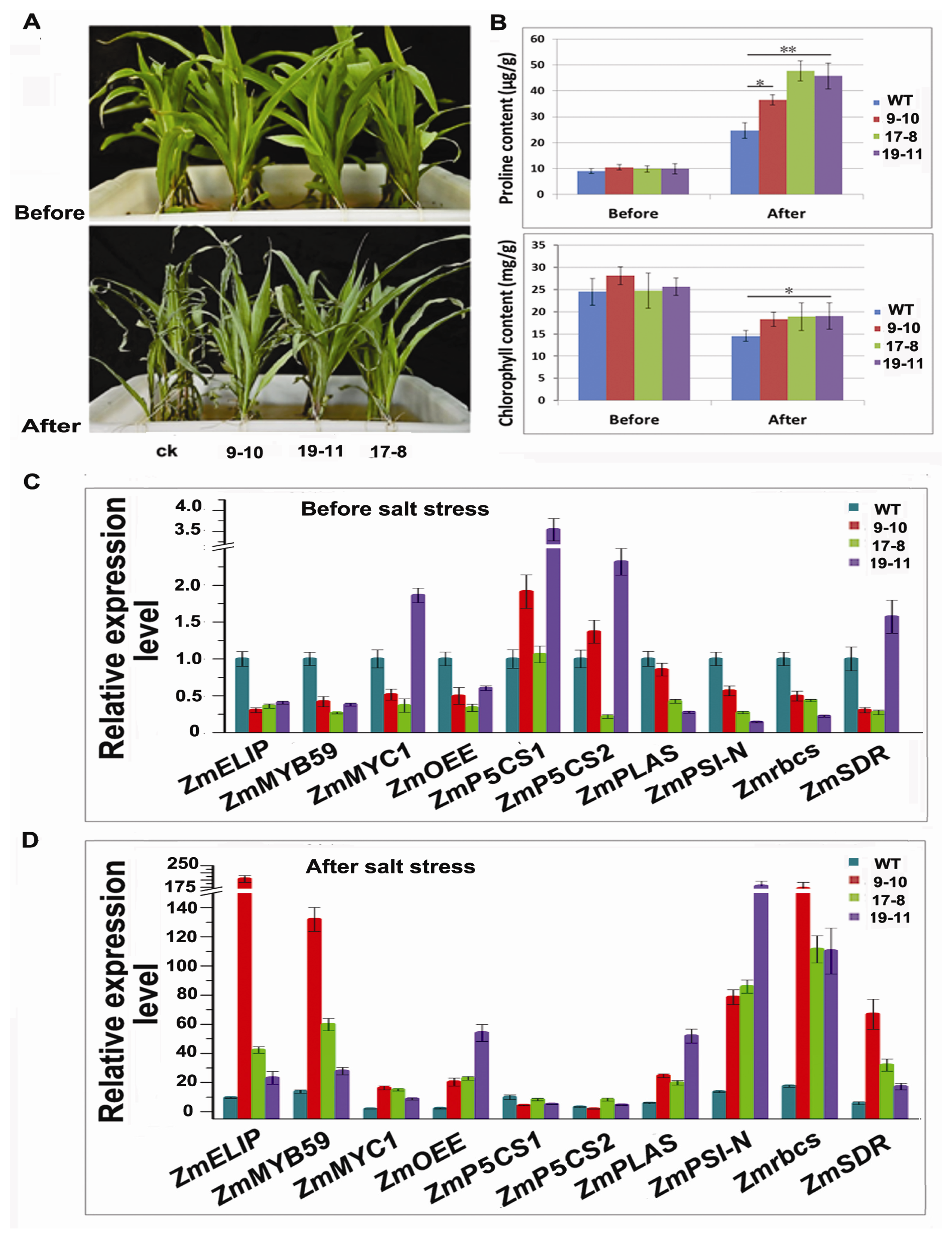
Figure 5A. Before treatment, these transgenic lines displayed similar growth situation as non-transformed control, suggesting that the expression and insertion of
TSRF1 into maize genome did not influence the growth of transgenic lines. After treated in Hoagland solution containing 200 mM NaCl for 48 h, both non-transformed and transgenic plant leaves displayed wilting and losing green symptoms. However, the degree of control seedlings was much more serious than that of transgenic lines. All the leaves of control seedlings were wilting and rolling. However, the first and second new leaves were still flat and green in transgenic seedlings.
Proline was considered to be an osmoprotectant to protect plants against the damage resulting from abiotic stress [
53]. Plants accumulate high level of proline under stress conditions to prevent the detrimental change. In addition, there is strong evidence that salt stress affects photosynthetic enzymes, chlorophyll and carotenoids in plants [
54]. Therefore, we measured the free proline and chlorophyll content change in control and transgenic lines after salt treatment. As it is shown in
Figure 5B, the free proline contents in control and transgenic seedlings were at similar level before treatment and increased dramatically after salt treatment. However, the proline content in the three transgenic lines increased at a much larger extent. Similarly, the chlorophyll content in transgenic seedlings decreased at a lower extent after salt treatment, indicating the more tolerance of
TSRF1 overexpressing transgenic maize.
It was previously shown that the constitutive expression of
TSRF1 enhanced rice salt and drought resistance by activating the expression of MYB, MYC, proline synthesis and photosynthesis-related genes [
32]. Therefore, we further investigated whether TSRF1 affects the expression of these genes in maize. Ten genes were selected to be analyzed, and the results were shown in
Figure 5C,D. The expressions of these genes were found to be different between transgenic lines. This might be due to the different transgenic lines with different target gene insertion sites and the expression levels of
TSRF1. Under normal conditions, the expression level of
ZmMYC1 in line 19–11,
ZmP5CS1 (Δ
1-pyrroline-5-carboxylate synthetase 1) and
ZmP5CS2 (Δ
1-pyrroline-5-carboxylate synthetase 2) in line 9–10 and 19–11 and
ZmSDR (short-chain dehydrogenase/reductase) in line 19–11 were increased, while the expression of these four genes in other lines were inhibited. The other six genes, including stress responsive transcription factor genes—
ZmMYB59—photosynthesis-related genes—
ZmELIP (low molecular mass early light-inducible protein HV90),
ZmPSI-N (photosystem I reaction center subunit N),
ZmOEE (photosystem II oxygen-evolving complex protein),
Zmrbcs (encoding ribulose 1, 5-bisphosphate carboxylase small subunit) and
ZmPLAS (encoding plastocyanin)—were inhibited in all these three transgenic lines, which was different from that in rice [
32]. This indicated that TSRF1 was probably involved in different regulatory pathways in maize from that in rice. Furthermore, we detected the expression changes of these genes in wild-type and transgenic lines after salt stress. As is shown in
Figure 5C, the expression of all these ten genes were increased by 2–16-fold in wild-type after salt treatment, indicating the endogenous regulation of salt responsive genes. Intriguingly, much more fold accumulations of these genes were induced after salt stress in
TSRF1 overexpressing lines. For example, the expression of
Zmrbcs had increased as high as 171-, 111- and 110-fold in transgenic lines 9–10, 17–8 and 19–11, respectively, compared with wild-type. The sharp induction of photosynthesis-related genes (
ZmELIP,
Zmrbcs and
ZmPSI-N) was consistent with the result of chlorophyll content change in
Figure 5B. TSRF1 increases ABA sensitivity in tobacco [
31,
55] and rice [
32].
SDR, an ABA biosynthesis related gene, was increased in
TSRF1 overexpressing rice [
32]. On the contrary, it was slightly suppressed in line 9–10 and 17–8 under normal condition. After salt treatment, surprisingly, TSRF1 triggered significant
ZmSDR induction. All these results suggested the complicated interaction of TSRF1 and ABA in plants. The accumulation of transcription factor gene
ZmMYB59 increased much more than
ZmMYC1 in all the transgenic lines, demonstrating the different roles of stress responsive transcription factors in salt stress. They may induce different other downstream salt related genes. These results indicated the complexity mechanism of
TSRF1 that involved in maize salt stress regulatory network.
Moreover, we noted that the expression levels of
ZmP5CS1 and
ZmP5CS2 were not or slightly increased in transgenic lines under salt condition, compared to wild-type, even though high level of proline accumulation was observed (
Figure 5B,C). It is reported that the activity of P5CS, a key enzyme representing a rate-limiting step in proline biosynthesis, is regulated through feedback inhibition by proline [
56–
59]. It is possible that a large amount of proline is synthesized at the early stage of salt stress and maintained this high level throughout the salty condition. Therefore, high activity of P5CS or high level of
P5CS transcript might be not necessary for high proline content maintaining under salt condition.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}