1. Introduction
Nitrogen (N) is one of the essential macronutrients required for plant growth and development and thus is often a major limiting factor for plant productivity and crop yield [
1,
2]. Nitrogen is not only a constituent of key cell molecules, such as amino acids, nucleic acids, chlorophyll, ATP and several plant hormones, but is also the pivotal regulator of numerous biological processes, including carbon metabolism, amino acid metabolism and protein synthesis [
3,
4]. Higher plants can absorb and use various forms of nitrogen components from the soil, most notably the inorganic ions ammonium (NH
4+) and nitrate (NO
3−). These ions are believed to be the principal nitrogen sources for plant growth in agricultural and most natural environments, as it is required in greater amounts than any other mineral nutrient [
5]. Because ammonium assimilation requires less energy than nitrate, ammonium is the preferential nitrogen source for root uptake, particularly in nitrogen-deficient plants [
6,
7]. Carbon (C) is also essential for plants to keep their fundamental growth and development. Various carbohydrates, for example, glucose, sucrose and organic acids provide both the energy and the carbon skeletons for NH
4+ assimilation to produce amino acids and proteins. The proteins, in particular enzymes, are essential for nearly all cellular activities. Several studies have emphasized the importance of the coordination of carbon and nitrogen metabolism [
8,
9,
10,
11,
12]. The optimal functioning of the metabolic pathways for carbon and nitrogen assimilation in plants, and maintaining an appropriate balance or ratio of carbohydrates to nitrogen metabolites in the cell, which is referred to as the “carbon/nitrogen balance”, are critical for determining plant growth, biomass accumulation and seed production [
8,
9,
10,
11,
12].
Rice (
Oryza sativa), a staple food for over half of the world’s population, is one of the most important crops worldwide. NH
4+ is the major available form of nitrogen that is used for growing rice plants in paddy fields and requires ammonium transporters (AMTs) in the root plasma membrane, which belong to the AMT/MEP/Rh (Ammonium Transporter/Methylamine Permease/Rhesus) protein superfamily with homologs in bacteria and fungi [
13,
14]. Several ammonia transporters, including bacterial AMT; plant AMT; and human RhAG, RhBG, and RhCG, have been suggested to support electroneutral NH
3 transport, NH
3/H
+ symport, NH
4+ transport, NH
4+/H
+ antiport and NH
4+/H
+ symport [
15,
16,
17,
18,
19,
20,
21,
22,
23,
24,
25]. AMT/MEP proteins are integral membrane proteins that harbor 11 transmembrane helices and intracellular
C terminals [
26,
27]. Biochemical studies have revealed that plant AMTs form trimers [
16]. Recently, a different membrane protein type, aquaporins from the MIP (Major Intrinsic Proteins) superfamily, was also demonstrated to transport ammonia [
28].
The cloning and expression analysis of
AMT genes in several plant species have been reported to date.
Arabidopsis harbors six AMT-type ammonium transporters, which are encoded by five genes (
AtAMT1-1,
AtAMT1-2,
AtAMT1-3,
AtAMT1-5,
AtAMT2-1) expressed in the roots and one gene (
AtAMT1-4) expressed in the pollen [
29].
AtAMT1-1 and
AtAMT1-3 are expressed in rhizodermal and cortical cells, where they confer high-capacity and high-affinity ammonium uptake [
30,
31]. In addition, reciprocal leaf and root expression of
AtAMT1-1 in response to nitrogen starvation has been reported by Engineer and Kranz [
32].
AtAMT1-2 is expressed in endodermal and cortical cells and most likely plays a major role in the uptake and retrieval of ammonium from the apoplast in the root with a lower-affinity [
31]. However,
AtAMT2-1 is more highly expressed in shoots relative to roots, and no ammonium influx activity has been observed in the root [
31,
33,
34].
AtAMT1-5 is expressed in rhizodermal root cells and contributes 5%–10% to the overall ammonium uptake capacity [
31].
AtAMT1-4 is a pollen-specific high-affinity ammonium transporter in the plasma membrane [
29]. Rice roots harbor four families of
AMT genes:
OsAMT1,
OsAMT2,
OsAMT3 and
OsAMT4.
OsAMT1 members have been characterized as a high-affinity transport system (HAT), share a highly similar sequence to each other, while the other three families have been characterized as a low-affinity transport system (LAT) [
13,
35]. Sonoda
et al. [
35] demonstrated distinct expression patterns in the
OsAMT1 gene family.
OsAMT1-1 is constitutively expressed in the shoots but is stimulated by ammonium in the roots;
OsAMT1-2 expression is root-specific and ammonium-inducible, whereas
OsAMT1-3 is also expressed specifically in the roots but is repressed by nitrogen.
OsAMT2-1 encodes an ammonium transporter, which is more closely related to the yeast MEP transporter sequence and is constitutively expressed in the shoots and roots [
36,
37]. Several studies revealed a feedback regulation and a distinct nitrogen-dependent regulation for the rice
AMT genes, which differs from that in tomato or
Arabidopsis. Sonoda
et al. identified cytosolic glutamine as a promising regulatory factor of the
OsAMT1 genes [
35]. Additionally,
AMT genes in several other plant species have been identified and characterized, including
Brassica napus (
BnAMT1-2) [
38],
Lotus japonicas (
LjAMT1-1,
LjAMT2-1) [
39,
40] and
Lycopersicon esculentum (
LeAMT1-1,
LeAMT1-2 and
LeAMT1-3) [
41,
42,
43].
Generally, the expression of the AMT1 isolated from different plant species are transcriptionally regulated. However, ectopically expressing of the
AtAMT1-1 gene under the control of a 35S promoter in transgenic tobacco plants revealed that the
AtAMT1-1 gene can also be post-transcriptionally regulated [
44]. A post-translational regulation of ammonium transport activity has also been described for the AMT1 proteins in
Arabidopsis. Loqué
et al. reported that the cytosolic trans-activation domain in AMT was essential for ammonium uptake, a mutation in the cytosolic AMT1-1
C terminus attenuated the strict dependence on allosteric trans-activation [
45,
46]. The oligomerization of plant AMTs is critical for allosteric regulation of transport activity, in which the conserved cytosolic
C terminus functions as a trans-activator [
47]. AMT1-1 and AMT1-2 are allosterically regulated by
C-terminal phosphorylation, which trans-inhibited the activation of AMT1 subunits in a trimeric complex [
45,
48,
49]. AMT1-3 harbors a phosphomimic residue in its
C terminus regulates both homo- and hetero-trimers in a dominant-negative fashion
in vivo [
47].
C-terminal phosphorylation of AMT1 is rapidly triggered by an external ammonium supply and decreases ammonium uptake by the roots and thereby quickly inactivating transport in a potentially toxic environment [
49]. Additionally, Graff
et al. [
50] also reported that
N-terminal cysteines affected the oligomer stability of the allosterically regulated ammonium transporter LeAMT1-1.
As we known, in agriculture, rice growth and yield requires abundant N. In order to meet the high production of rice, large amounts of synthetic N fertilizers are applied on arable land by farmers. However, crop plants use less than half of the applied N fertilizers [
51]. Zhang
et al. [
52] reported that the N use efficiency of midseason rice in China is less than 30%. The applications of large quantities of synthetic N fertilizers to increase crop yield are not economically sustainable and placed a heavy economic burden on farmers, and also result in environmental pollutions. Because of the high-affinity NH
4+ uptake function of
AMT1-3 in rice, it is a good candidate gene for use in transformation strategies aimed at improving nitrogen use efficiency and rice yield. However, no study has been reported about the function of
AMT1-3 gene in rice growth and carbon-nitrogen metabolism to date. In order to evaluate the effect of
AMT1-3 gene on rice growth, nitrogen absorption and metabolism, we generated
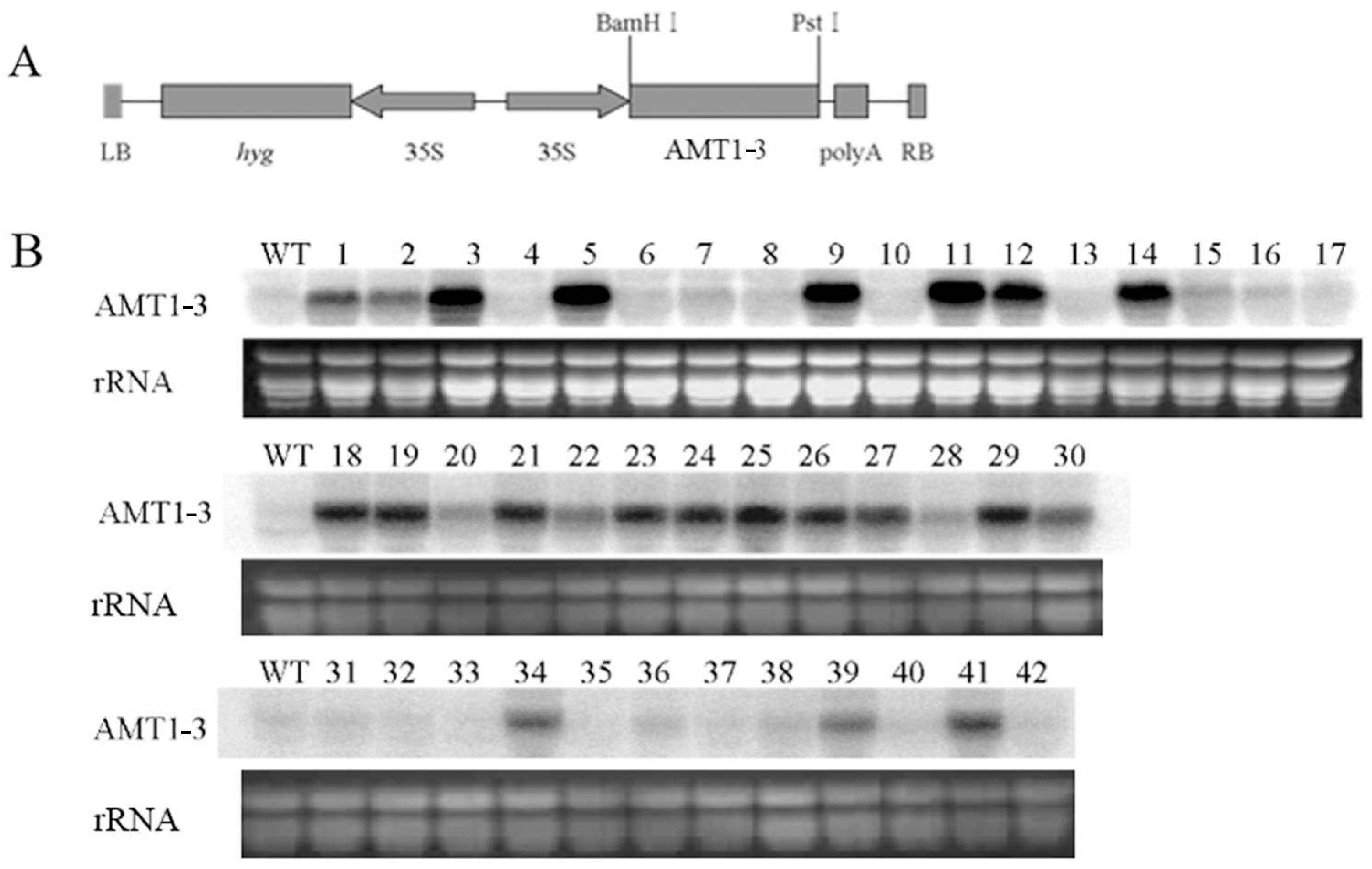
AMT1-3-overexpressing plants
via the
Agrobacterium-mediated transformation method using the
CaMV35S promoter in this study, and analyzed the growth phenotype, yield, leaf SPAD (Soil and Plant Analyzer Development) value, photosynthesis, carbon and nitrogen metabolic status and gene expression profiles under four different N levels (0× N, 0.1× N, 1× N and 5× N) at both tillering and heading stages. Results revealed that the overexpressing of
AMT1-3 gene altered rice growth and development, yield, C/N ratio, soluble proteins and carbohydrates, carbon and nitrogen metabolites, gene expression patterns in transgenic plants when compared to the wild-type plants. Moreover,
AMT1-3 may act as a signal sensor to regulate plant growth in addition to its function in NH
4+ uptake in the root. Our results provided a new understanding of the function of
AMT1-3 genes in rice growth, yield, carbon and nitrogen metabolism, and a reference for the molecular breeding of the high nitrogen use efficiency rice cultivar using transgenic strategy.
3. Discussion
As
OsAMT1 members, including
OsAMT1-1,
OsAMT1-2 and
OsAMT1-3, have been characterized as high-affinity NH
4+ transport genes in rice [
13,
35], we expected to obtain
OsAMT1-overexpressing transgenic rice plants with higher ammonium uptake capacity, improved nitrogen use efficiency and a low nitrogen tolerance using the
Agrobacterium-mediated transformation method. Although we generated
AMT1-2-,
AMT1-3-overexpressing plants, we failed to obtain
AMT1-1-overexpressing plants. Unexpectedly,
AMT1-3-overexpressing plants displayed poor growth, but no significant differences between
AMT1-2-overexpressing plants and wild-type plants were observed. In order to understand why poor growth phenotype exhibited in
AMT1-3-overexpressing transgenic rice plants and evaluate the effect of
AMT1-3 gene on the rice growth, nitrogen absorption and metabolism, we analyzed the growth phenotype, yield, carbon and nitrogen metabolic status and gene expression profiles of the
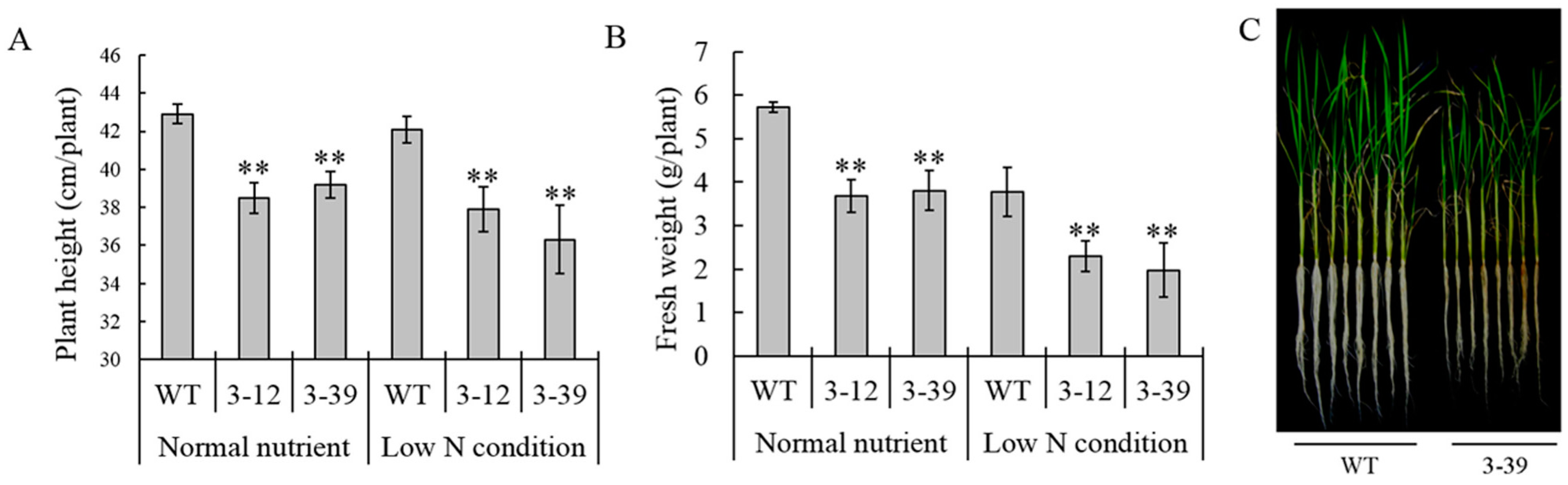
OsAMT1-3-overexpressing rice and wild-type Zhonghua 11 under four different N levels (0× N, 0.1× N, 1× N and 5× N) at both tillering and heading stages in detail. Results showed that despite the higher
AMT1-3 mRNA levels in transgenic plants, the root length, plant height, root and shoot biomass and yield were significantly decreased in
AMT1-3-overexpressing plants compared with wild-type plants (
Figure 2 and
Figure 3;
Table 2). The contents of total carbon and nitrogen, soluble proteins and carbohydrates, individual metabolites (such as sugars, organic acids, amino acids), the expression patterns of key genes involved in carbon and nitrogen metabolic pathway changed significantly in
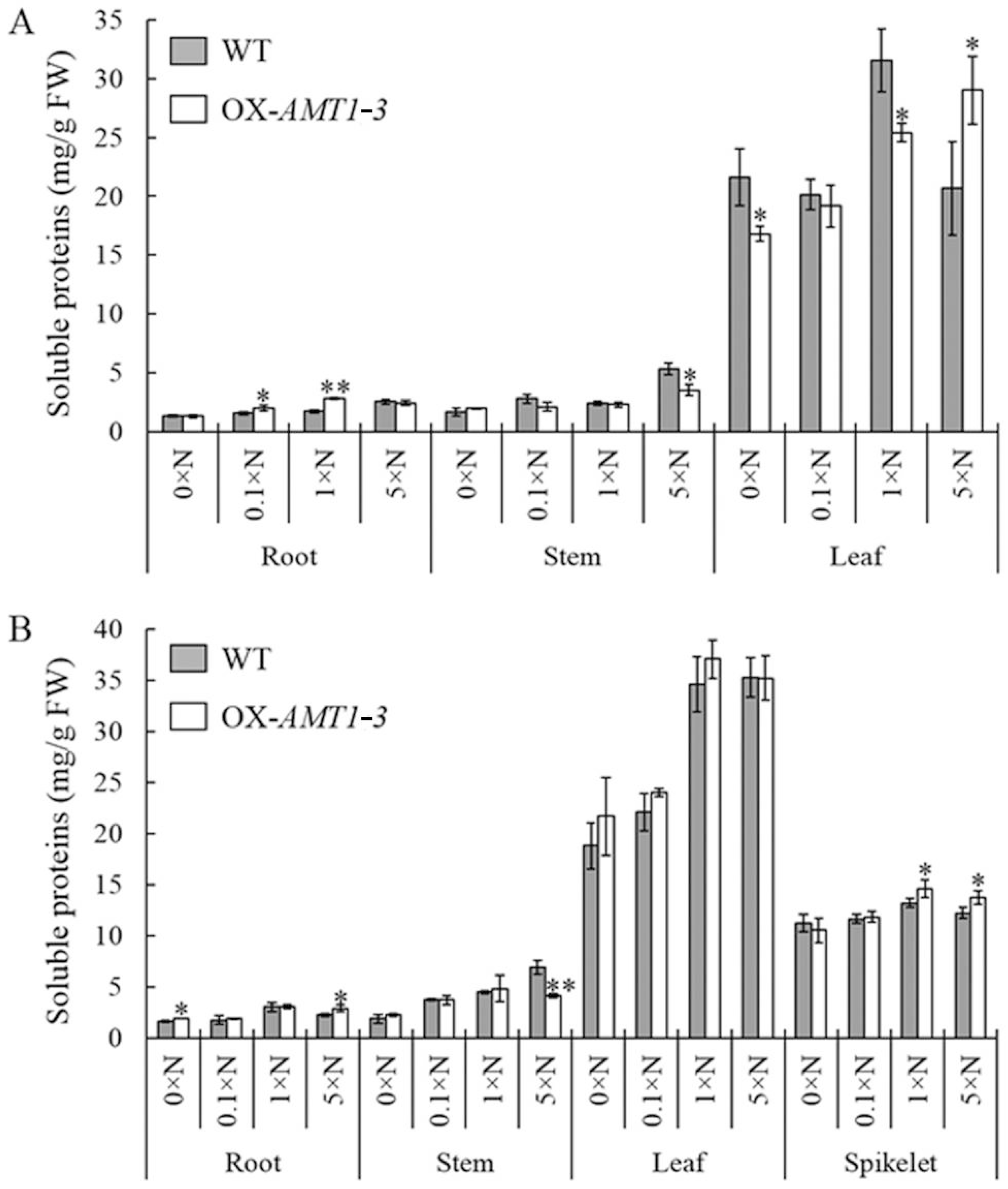
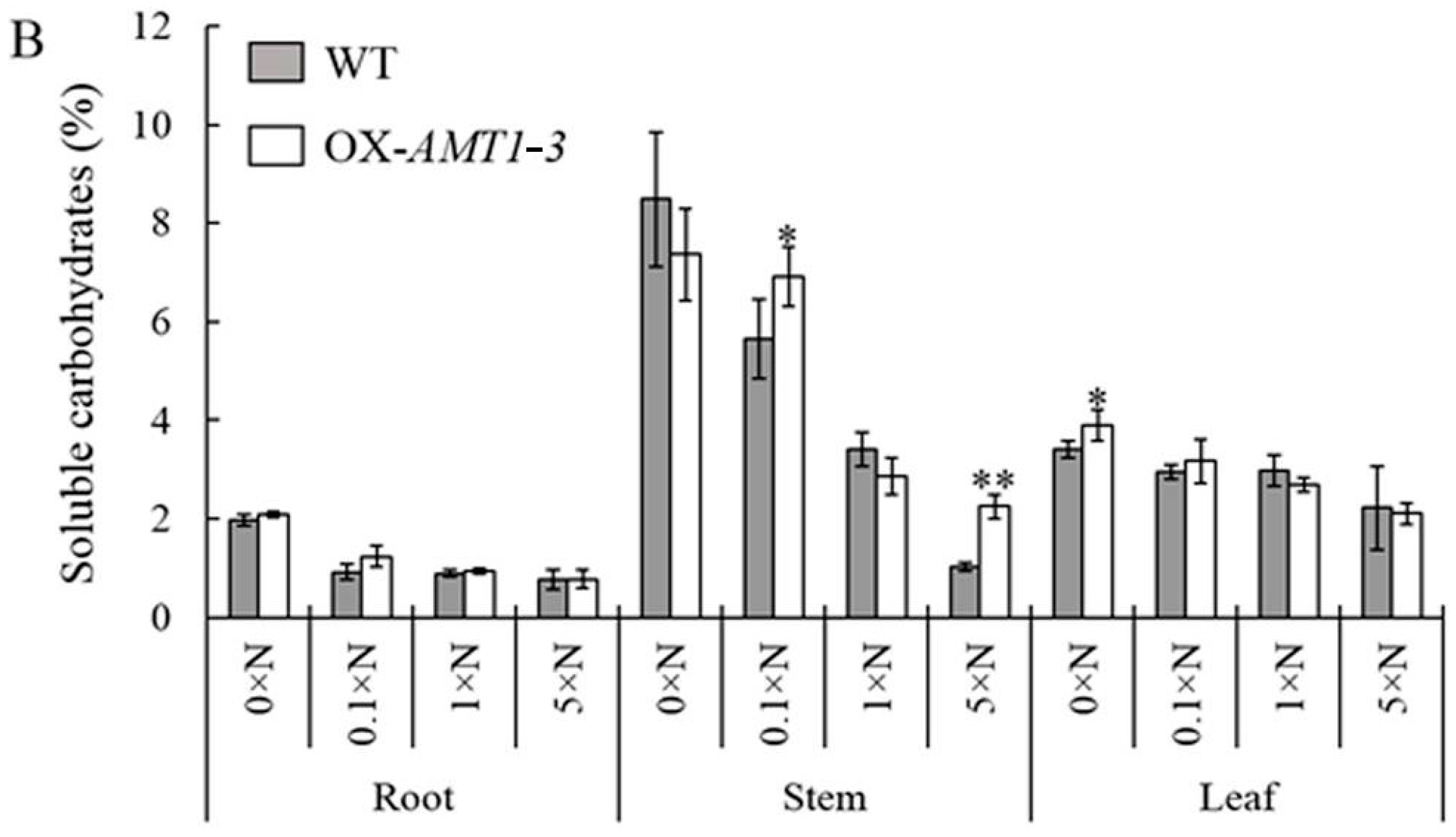
AMT1-3-overexpressing plants (
Figure 5,
Figure 6,
Figure 7 and
Figure 8;
Table 3). The discordant of carbon and nitrogen metabolic status in transgenic plants caused by the
OsAMT1-3 overexpressing may be the main reason for poor growth and yield in our investigation. Similar results were reported that overexpressing of
OsAMT1-1 gene caused decreased shoot and root biomass in transgenic lines during seedling and early vegetative stage, especially when grown under high ammonium nutrition [
53,
54]. However, completely different results were reported by Ranathunge
et al. [
55] that transgenic rice lines overexpressing the
OsAMT1-1 gene had higher plant growth rate and grain yield, especially under suboptimal NH
4+ levels.
As we known, both carbon and nitrogen nutrients are essential for various cellular functions. Recently, cellular carbon and nitrogen metabolism was demonstrated to be tightly coordinated [
8,
9,
10,
11,
56]. In addition to independent utilization, the coordination and optimal functioning of the metabolic pathways governing nitrogen and carbon assimilation in plants were demonstrated to be critical for plant growth and, ultimately, biomass accumulation and yield production [
10,
56]. Maintaining an appropriate balance or carbohydrate to nitrogen metabolite ratio in the cell, which is referred to as the “carbon/nitrogen balance”, is extremely important for regulating plant growth, development and production yield [
8,
9,
11]. In our study, we found 4.6%–9.9% and 4.7%–15.5% decreases in
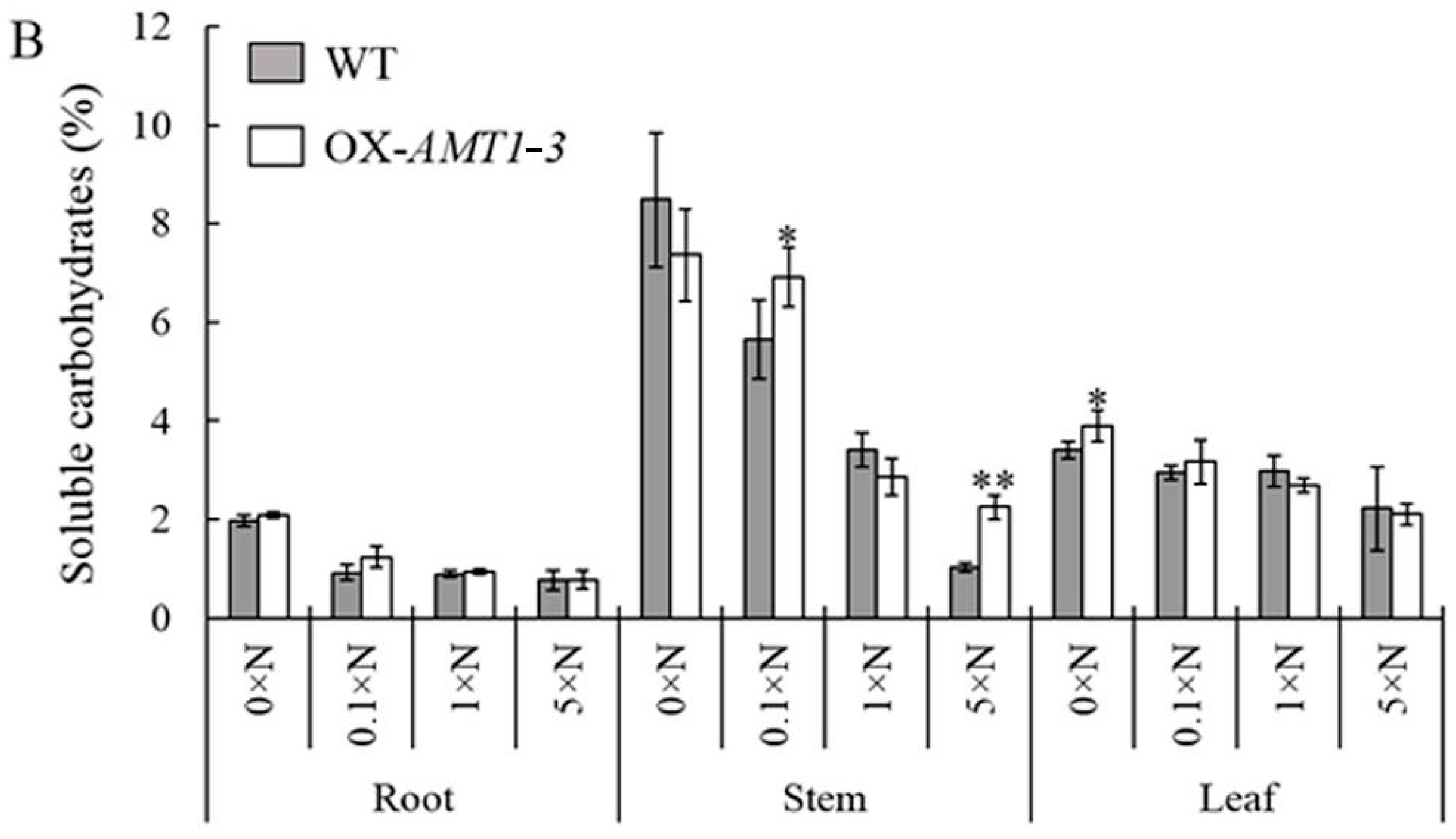
AMT1-3-overexpressing plant leaf C and N content, respectively, which resulted in a higher (9.4%–11.7% increases) leaf C/N ratio in
AMT1-3-overexpressing plant leaves (
Table 3). Additionally, distinct changes between the soluble protein and carbohydrate concentrations were observed in
AMT1-3-overexpressing plants (
Figure 5 and
Figure 6). Therefore, combined with our results, we hypothesized that the carbon and nitrogen metabolic imbalance caused by
AMT1-3 overexpressing attributed to the poor growth and yield of transgenic plants.
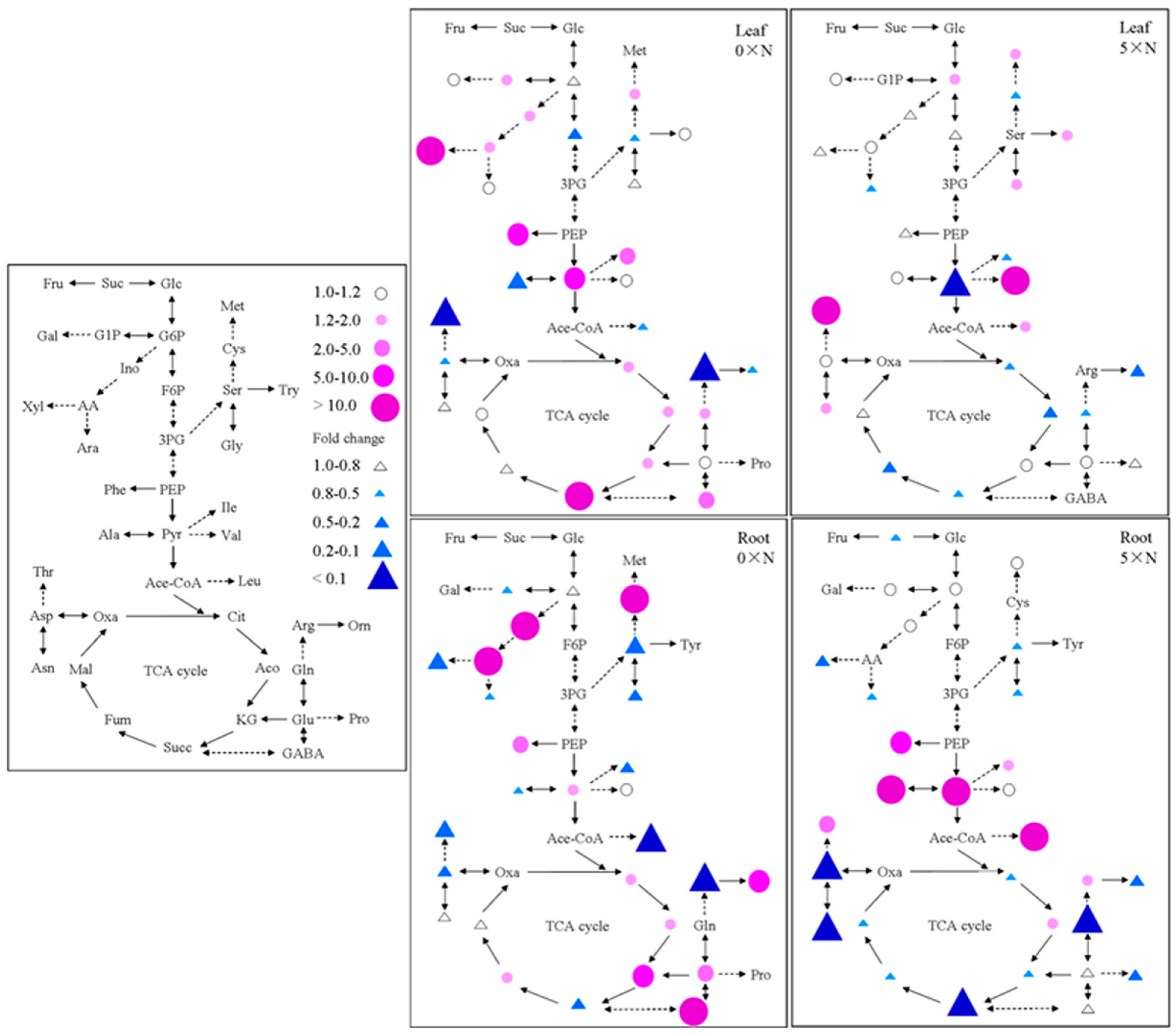
To further study the impact of higher
AMT1-3 mRNA levels at both the physiological and molecular level, we analyzed the individual carbon and nitrogen metabolites and the gene expression levels in the roots and leaves of
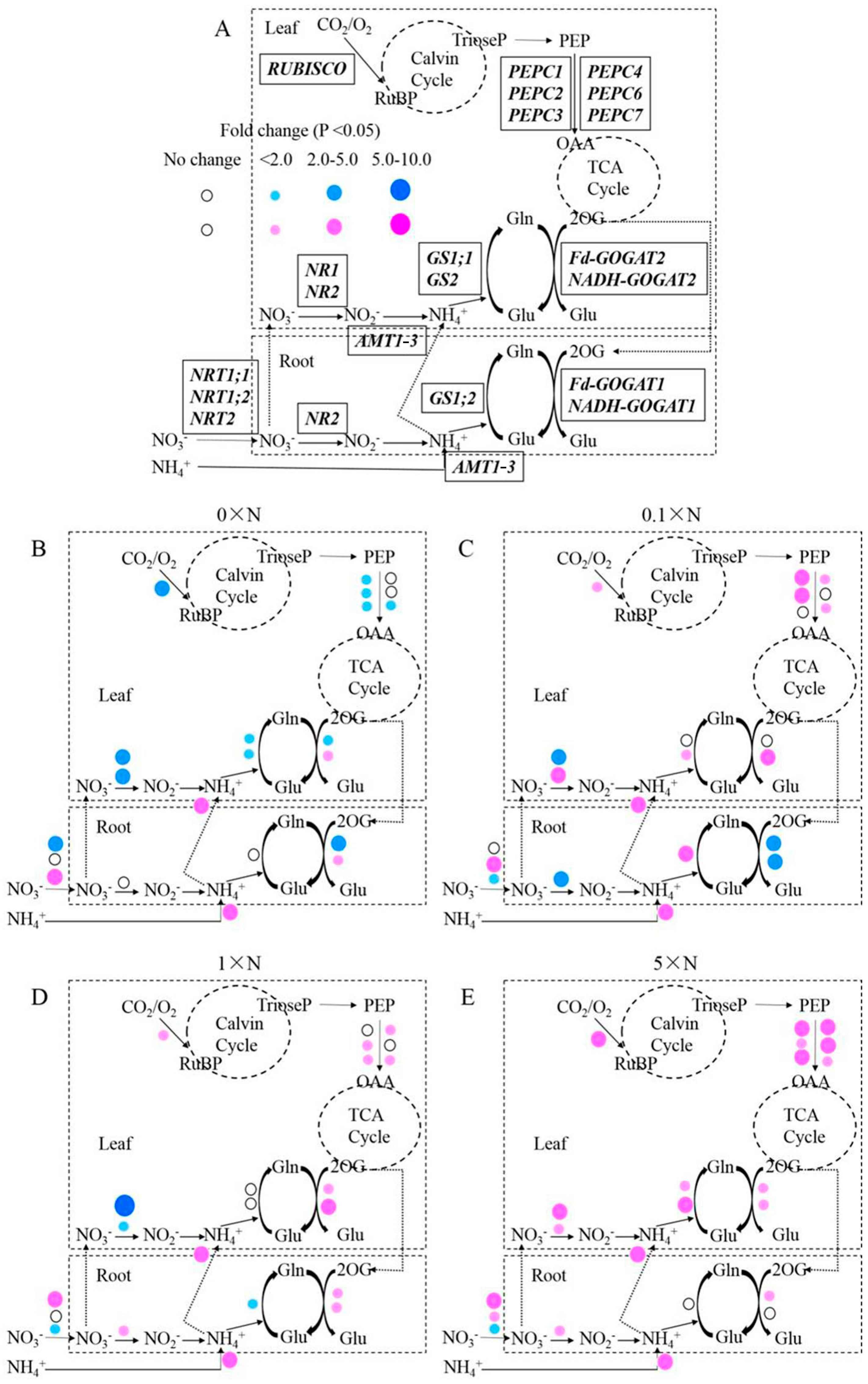
AMT1-3-overexpressing plants and wild-type plants. Consistent results of the metabolite concentrations and gene expression levels were showed in this study. Under the 0× N growth condition, given an almost complete lack of nitrogen (a residual amount of nitrogen may reside in water) in the nutrient solution, the nitrogen uptake, reduction and assimilation levels declined in leaves. The expression level of
NRT1;1, which encodes a low-affinity NO
3− transporter [
57], was repressed, whereas
NRT2, which encodes a high-affinity NO
3− transporter [
58], was induced in the root. Although the
AMT1-3 gene was overexpressing under the 35S promoter, the NH
4+ concentration in roots and leaves significantly decreased. Together with the lower NO
3− uptake and transport by NRTs in roots and leaves, the expression levels of the genes (
NR1,
NR2,
GS1;1,
GS2,
Fd-GOGAT1 and
Fd-GOGAT2) involved in NO
3− reduction and NH
4+ assimilation were significantly decreased, which resulted in reduced total free amino acids and soluble proteins (
Figure 5,
Figure 7 and
Figure 8). During NH
4+ assimilation, numerous organic acids provided carbon skeletons to produce a variety of amino acids [
59]; therefore, an accumulation of total organic acids may have been observed because of the decreased nitrogen assimilation status in leaves of
AMT1-3-overexpressing plants (
Figure 8). The expression levels of
RUBISCO,
PEPC1,
PEPC2,
PEPC3 and
PEPC7 were decreased, which lead to a decline in total sugars and soluble carbohydrates in leaves of
AMT1-3-overexpressing plants (
Figure 6,
Figure 7 and
Figure 8). Under the 5× N growth condition, given the much higher nitrogen content in the nutrient solution, higher levels of nitrogen uptake, reduction and assimilation were observed in both roots and leaves of
AMT1-3-overexpressing plants. The expression of low-affinity NO
3− transporter genes (
NRT1;1 and
NRT1;2) [
57] were induced, whereas the expression of the high-affinity NO
3− transporter gene
NRT2 [
58] was repressed in the root, which resulted in more NO
3− absorption and higher expression of genes (
NR1,
NR2,
GS1;1,
GS2,
Fd-GOGAT1,
Fd-GOGAT2 and
NADH-GOGAT2) involved in NO
3− reduction and NH
4+ assimilation. As a result, total free amino acids and soluble proteins accumulated in leaves of
AMT1-3-overexpressing plants, which depleted total organic acids to provide carbon skeletons for amino acid production. This large consume of organic acids promoted the breakdown of sugars. Conversely, the expression levels of
RUBISCO,
PEPC1,
PEPC2,
PEPC3,
PEPC4,
PEPC6 and
PEPC7 dramatically increased in leaves of
AMT1-3-overexpressing plants to produce more carbohydrates (
Figure 7).
Plants have evolved a complex profile of responses to cope with changes in soil nitrogen availability, which is mediated by a stringent control of expression and/or activity of proteins involved in nitrogen transport and assimilation [
59]. Recent studies on nutrient effects in plants have focused on their potential roles as signaling molecules in addition to their roles as building blocks of organic matter or cofactors [
60]. Nitrogen sensing appears to regulate a variety of physiological and developmental processes in plants [
8,
61]. For example, NO
3− is a positive signal required for the induction of NO
3− uptake and its reduction, and the metabolized products of NO
3−, NH
4+ and its assimilation products Glu and Gln are believed to exert negative effects on NO
3− uptake and reduction [
35,
62,
63,
64,
65]. Bacterial and fungal AMTs have been demonstrated to act as transceptors with dual functions as ammonium transporters and receptors that mediate ammonium-triggered changes in morphology or the transcription of target genes [
66]. Plant AMTs may also be transceptors. For example, AMT1-3 can regulate lateral root branching in response to localized ammonium supplies [
67]. More recent reports have demonstrated that
OsAMT1-3 is expressed specifically in roots but repressed by nitrogen, which indicates that
OsAMT1-3 participates not only in ammonium uptake but also in ammonium sensing in rice [
14,
35]. In this study, we expected to enhance ammonium uptake capacity in roots and to improve nitrogen fertilizer use efficiency and yield formation by overexpressing the
AMT1-3 gene in rice. In contrast, our transgenic plants grew poorly and displayed a low yield production and an unbalanced carbon and nitrogen metabolic status (
Figure 3;
Table 2 and
Table 3). We hypothesized that the
AMT1-3 transcriptional level may act as a signal sensor to regulate carbon and nitrogen metabolism in rice.
AMT1-3 is a high-affinity NH
4+ transporter gene, its transcriptional level is mainly expressed in rice roots under low nitrogen in the environment and repressed with nitrogen supplementation [
13,
35] The high transcriptional level of
AMT1-3 in our transgenic rice may mimic a nitrogen starvation signal in the environment and thus retard plant growth and development to ensure the plant can complete its entire life cycle. Recently, Gaur
et al. validated
OsAMT1-3 as a biomarker for detecting available nitrogen pools both inside cells and in the soils around the root [
14]. Similar results were reported by Yuan
et al. who attempted to increase the ammonium uptake capacity via ectopic expression of
AMTs; however, this largely failed because their transport capacities are tightly regulated by ammonium [
31]. In addition, excess ammonium intake deregulates cellular pH homeostasis and primary metabolism [
68]. Therefore, plant roots repress ammonium uptake at elevated ammonium supplementation [
49,
64]. Furthermore, several published works have demonstrated that
C-terminal phosphorylation can mediate intermonomeric trans-inhibition of ammonium transport in plant roots and that the allosteric regulation of transporter activities even extends to heteromeric AMT protein complexes in
Arabidopsis [
47,
49]. Additionally,
N-terminal cysteines were demonstrated to affect oligomer stability of the allosterically regulated ammonium transporter LeAMT1-1 [
50]. Thence, the post-transcriptional and post-translational regulation of plant AMT cannot be ignored in further studies.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





