1. Introduction
Sacha inchi (
Plukenetia volubilis L.), a member of the Euphorbiaceae, is native to the rain forest of South America [
1,
2]. Because its seed oil is rich in polyunsaturated fatty acids (PUFAs) and lipovitamins, Sacha inchi has great potential economic value to the food and pharmaceutical industries [
3,
4]. Moreover, Sacha inchi oil is also a good feedstock for biodiesel production [
5]. To promote gene function studies in Sacha inchi, transcriptomic analysis has been performed for the period of seed development, and numerous key genes involved in the regulation of seed oil biosynthesis have been identified [
6]. A good characterization of expression profiles of these key genes will facilitate a better understanding of gene function in seed oil biosynthesis.
Characterized by high sensitivity, specificity and accuracy, real-time quantitative PCR (RT-qPCR) has become the preferred method for detecting and measuring gene expression [
7,
8,
9]. A prerequisite for the reliable analysis of gene expression is the normalization of RT-qPCR data, which can minimize the non-specific variations caused by variations in the quantity and quality of mRNA and variations in the efficiencies of reverse transcription and PCR [
10,
11,
12,
13]. Therefore, the selection of appropriate reference genes as internal controls that are expressed at constant levels among tissues and over time is very important.
In the last decade, several statistical algorithms have been developed for the selection of reference genes for RT-qPCR analysis, such as the delta cycle threshold (Δ
Ct) [
14], geNorm [
15], BestKeeper [
16] or NormFinder [
17] algorithms. The Δ
Ct method ranks the candidate genes by comparing the relative expression of pairwise under a given set of experimental conditions [
14]. The Δ
Ct method indicated the mean of standard deviation (SD) of each candidate reference genes, and the candidate with the lowest SD value was proposed to be the most stable gene [
14]. The geNorm is a Visual Basic application tool that relies on the principle that the expression ratio of two perfect reference genes should be constant under different development stages or in various plant tissues. The expression stability (M) is calculated based on the average pairwise variation between all reference genes tested. The gene with a lower M value indicated the gene expression is more stable [
15]. The BestKeeper program evaluates the most stably expressed genes based on the coefficient of variance (CV) and SD of the quantification cycle (Cq) values. The lower coefficient of variance and standard deviation (CV ± SD) indicated the gene expression was more stable [
16]. The NormFinder program is based on a variance estimation approach, which ranks the candidate genes according to the stability of a gene under a given set of experimental conditions compared to the rest of the tested genes. The more stably expressed genes are indicated by the lower average expression stability values (M values) [
17]. The application of these algorithms has simplified the identification of reliable reference genes by enabling the rapid calculation of the expression stability and the determination of the optimal number of reference genes required for normalization [
18,
19].
The identification of optimal reference genes for RT-qPCR has been reported for several plants, including bamboo [
20],
Jatropha curcas [
21], coffee [
22], oil palm [
23], peach [
24] and
Petunia hybrida [
25]. However, a number of commonly used housekee** genes, such as
actin (
ACT),
elongation factor 1alpha (
EF1α),
glyceraldehyde-3-phosphate dehydrogenase (
GAPDH), and
ubiquitin(
UBQ), is insufficient for RT-qPCR normalization because of variations in expression in different species, tissues, developmental stages or environmental conditions. For sesame,
SiACT was recommended as the reference gene for seed development and germination, although
SiUBQ6 was better for bud development [
26]. For Chinese cabbage,
EF1α was reported to be the best reference gene among five tissues, and
GAPDH was most suitable for drought stress conditions [
27]. The
18S rRNA (
18S),
ACT and
GAPDH genes were reported to be expressed unstably in papaya (
Carica papaya) under numerous experimental conditions [
28]. Hence, the selection of multiple housekee** reference genes is required for the accurate normalization of gene expression levels under varied experimental conditions.
For this study, in order to reduce the likelihood that the reference genes exhibited regulated co-variation, a group of genes with varied roles in different cellular processes were chosen (
Table 1). The expression stabilities of twelve candidate reference genes (
18S,
ACT,
CYC,
EF1α,
GAPDH,
PLA,
RPII,
RPS13,
TEF2,
TUB,
UBL and
UCE) were examined in Sacha inchi seedlings and adult plants, during flower and seed development, and for the entire growth cycle of Sacha inchi. Our results indicate that traditional housekee** genes were less stably expressed than other reference genes in the given experimental datasets.
Table 1.
Selected candidate reference genes, primer sequences and PCR amplification characteristics.
Table 1.
Selected candidate reference genes, primer sequences and PCR amplification characteristics.
| Gene/GenBank Accession Number | Description | Function | Forward (F) and Reverse (R) Primer Sequences (5′→3′) | Amplicon Length | Tm (°C) | Amplification Efficiency (%) | Correlation Coefficient |
|---|
| 18S/KP729648 | 18S ribosomal RNA | ribosomal structure | F: ACCAGGTCCAGACATAGTAAGGATTGA | 140 bp | 81.73 | 106.40 | 0.999 |
| R: AGTTAGCAGGCTGAGGTCTCGTT |
| ACT/GADC01011038 | actin | cytoskeletal structural protein | F: CCAGAAGTCTTGTTCCAGCCATCTC | 185 bp | 80.66 | 105.78 | 0.999 |
| R: GCGGTGATCTCCTTGCTCATACG |
| CYC/GADC01018836 | cyclophilin | protein folding | F: GGCAAGATACGAACGGATCACAGTT | 145 bp | 82.95 | 108.93 | 0.999 |
| R: GGCACTCCACTCCGACTTCCTT |
| EF1α/GADC01006492 | elongation factor 1-alpha | protein biosynthesis | F: GGTATTCTCAAGCCTGGTATGGTTGT | 102 bp | 80.48 | 94.98 | 0.999 |
| R: GAGAGCCTCCTGAAGAGCCTCAT |
| GAPDH/GADC01052274 | glyceraldehyde-3-phosphate dehydrogenase | glucose metabolism | F: TGGCAAGCATATTCAGGCAGGAG | 116 bp | 81.63 | 94.98 | 0.999 |
| R: TTGGCTCATCAGGATTGTAGGTATCAG |
| PLA/KP729647 | phospholipase A22 | lipid catabolic process | F: ATACCATACAGAACGCAGCTTGTGAA | 101 bp | 79.92 | 103.33 | 0.998 |
| R: TTCCGCCAGTTCCAACCTATCCA |
| RPII/GADC01020629 | RNA polymerase II subunit | mRNA process | F: GCCTCGGTCTCATTCCTCTTACAAG | 109 bp | 82.44 | 104.17 | 0.999 |
| R: AACTCAACAGAACAATACTCGCACTGA |
| RPS13/GADC01008223 | 30S ribosomal protein S13 | DNA-templated transcription | F: TAATGCACAGCTTCCAGATGAC | 202 bp | 81.47 | 90.55 | 0.999 |
| R: AACCAGTCGCTTTGATTCTTCT |
| TEF2/GADC01000224 | transcription elongation factors-II | transcription | F: AGATTCAGAGCATGAAGAGGGAC | 182 bp | 82.18 | 104.17 | 0.996 |
| R: CGATCGGTATTTGTTGCGATTT |
| TUB/GADC01018931 | Tubulin beta-4 chain | structural constituent of cytoskeleton | F: ACAATTCACTGCCATGTTCAGGAGAA | 169 bp | 82.05 | 97.83 | 0.999 |
| R: GTCATCTTCGTAGTCACCTTCGTCATC |
| UBL/GADC01024109 | ubiquitin-like | protein binding | F: GCTACGTCTGCGTGGAGGAATG | 197 bp | 82.39 | 99.53 | 0.996 |
| R: TGTAGTCTGCCAATGTGCGTCC |
| UCE/GADC01034781 | ubiquitin-conjugating enzyme | ubiquitin-dependent protein catabolic process | F: TGGAATGGATGACGGAGACGACAT | 142 bp | 78.74 | 100 | 0.997 |
| R: AACACTTGGTGGCTTCTCTGGATAATC |
3. Discussion
As a result of its high sensitivity, specificity and cost-efficiency, RT-qPCR has greatly improved the quality of measurements of expression levels of target genes in biological samples [
32]. However, the accuracy of RT-qPCR analysis can be significantly affected by several factors, including RNA quality, the quantity of cDNA and the selection of reference genes [
9,
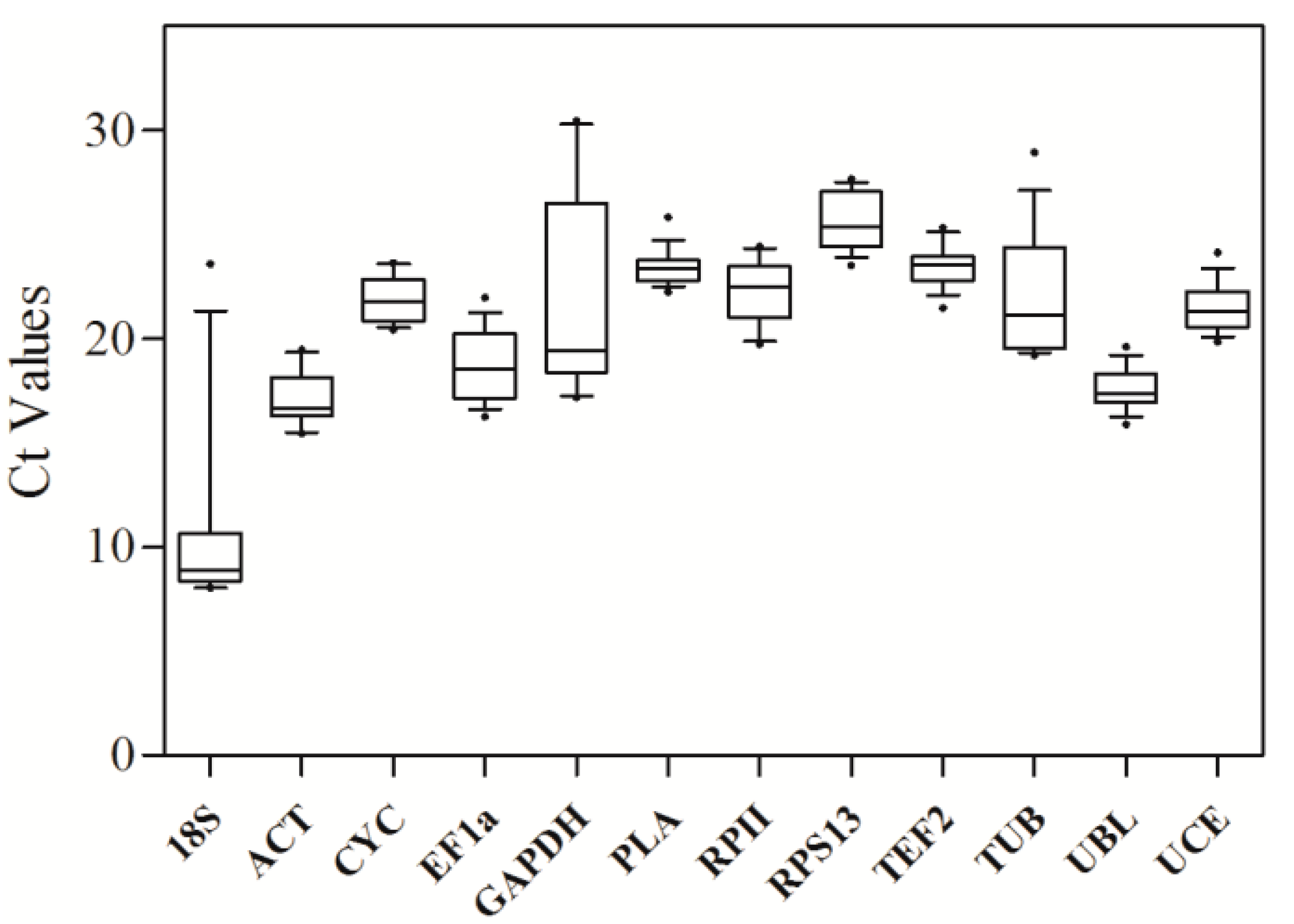
16]. To achieve high accuracy, a reference gene should have a relatively stable expression level in distinct biological samples, such as across tissues, developmental stages and experimental conditions. In this study, the expression stabilities of twelve candidate reference genes were estimated in various tissues and developmental stages of Sacha inchi. The
UCE,
ACT, and
PLA genes were found to be the most stable genes in seedlings. For roots, stems, leaves, flowers, and seeds from adult plants,
RPS13,
CYC, and
EF1α were recommended as reference genes for RT-qPCR. During the development of reproductive organs,
PLA,
ACT, and
UCE were the optimal reference genes for flower development, whereas
UCE,
RPS13, and
RPII were optimal for seed development.
In this study, four computational methods (Δ
Ct, BestKeeper, NormFinder and geNorm) were used to evaluate the stability of the expression levels of these twelve candidate reference genes. Here we found that the least stable genes computed by the four algorithms were almost the same, while the most stable genes differed. In the set of adult plant, the
18S gene was ranked last by all four algorithms, whereas the
RPS13, the
UCE, the
EF1α and the combination of
CYC and
RPS13 genes were ranked first by Δ
Ct, BestKeeper, Normfiner and geNorm, respectively (
Table 2). To obtain the most stable reference gene, we used the RefFinder tool that integrates the currently available major computational programs (Δ
Ct, BestKeeper, Normfinder and geNorm) to compare and rank the tested candidate reference genes. Based on the rankings from each above mentioned program, RefFinder assigned an appropriate weight to an individual gene and calculated the geometric mean of their weights for the overall final ranking. Accordingly, the
RPS13 was recommended as the most appropriate reference gene in adult plant (
Table 2).
The 18S ribosomal RNA is a component of the small subunit of eukaryotic ribosomes (40S). The
18S gene has been used as a reference gene for RT-qPCR normalization in many previous studies [
33,
34]. In
Jatropha, the
18S was applied to normalize the expression of
JcAOC and
JcBD1 in various tissues under salt and cold stress conditions [
35,
36]. However, in this study, the
18S gene was the least stable gene across three experimental datasets,
i.e., the adult plant, the seed developmental stage, and the entire growth cycle of Sacha inchi. The
18S gene has also been deemed inappropriate for gene expression analyses in
Pisum sativum [
37] and bamboo [
20]. The
GAPDH gene, which encodes a key enzyme involved in the glycolysis and gluconeogenesis [
38], is another commonly used reference gene. It is the most stable reference gene in
Jatropha over different tissues, developmental stages and experimental conditions [
21]. And it has been also recommended in flax [
39] and coffee [
22,
40]. However,
GAPDH has been reported as the least stable reference gene in oil palm [
23], peach [
24], Petunia hybrida [
25] and bamboo [
20]. Similarly, in the present study, the
GAPDH expression varied among tissues in Sacha inchi, except across the flower developmental stages in which it ranked the fourth. It is possible that
GAPDH is not only a key enzyme involved in glycolysis but also participates in other processes. The
TUB gene, which plays a crucial role in cell structural maintenance, has also been widely used as a reliable reference gene in switchgrass [
41] and peach [
24]. However, in our study,
TUB was identified as a poor reference gene, similar to results for potato [
42] and soybean [
43]. Taken together, these results indicate that the most stable reference genes differ among plants or tissues. Hence, the choice of reference genes is very important.
Our results indicate that
ACT is suitable for normalization in seedlings and during flower development in Sacha inchi. In
Jatropha,
ACT expression was more ubiquitous than in Sacha inchi, and was found across the different plant developmental stages and under cold-/drought-induced conditions [
21].
UCE was ranked among the top three most stable reference genes for all tissues, with the exception that it was ranked fourth for the adult plant. Therefore,
UCE is recommended for the normalization of gene expression in Sacha inchi.
UCE has also been identified as one of the most stable reference genes in switchgrass [
41], whereas
UCE was the most variable reference gene for the tung tree [
12] and
Jatropha curcas [
21]. In addition, we found that
CYC was ranked first for the entire growth cycle of Sacha inchi and second for the adult plant in this study. The
CYC gene was among the best reference genes for
Petunia hybrida [
37] and bamboo [
24]. The
RPS13 gene, which was used for the normalization of gene expression in
Petunia hybrida [
25], might also serve as a reliable reference gene for studies of adult plants, different developmental stages of seeds, and the entire growth cycle of Sacha inchi.
To illustrate the actual utility of validated reference genes in this study, the expression pattern of
PvoAG was examined in Sacha inchi.
AG belongs to the C-class genes in the ABC model of floral organ development [
44]. In
Arabidopsis,
AG was mainly expressed in inflorescences and flowers, and was involved in the regulation of stamen and pistil development [
44]. In poplar and strawberry,
AG was also highly expressed in flowers with low levels in leaves, stems and seeds [
36,
45]. Here, in Sacha inchi adult plants,
PvoAG was remarkably expressed in young inflorescences with relatively lower expression in seeds (90 DAP) when the most stable genes
RPS13 and
CYC were used for normalization (
Figure 3A). This result is similar to the
AG expressions in other plants mentioned above. However, when the least stable genes
GAPDH and
18S were applied, the expression level of
PvoAG was extremely high in seeds (90 DAP) (
Figure 3A). Thus, these results further proved the necessity of selection of reliable reference genes in gene expression studies.
To date, numerous studies have reported that when evaluating levels of target gene expression, the results are more pronounced and reliable when two or more reference genes are utilized [
42,
46,
47]. In this study, we have recommended the three most reliable reference genes for expression analyses of Sacha inchi for each of the aforementioned experimental conditions. The results of this study will help inform the selection of stable reference genes for future gene expression studies of Sacha inchi.




{kind=link}
{kind=link}
{kind=link}


