1. Introduction
Many plant-pathogen interactions are determined by the presence of resistance (
R) genes/alleles that enable plants to recognize pathogens and activate inducible defenses [
1]. Plant diseases have been reported to be caused by a wide variety of pathogens, such as
Fusarium sp. in fusarium head blight (FHB) [
2],
Sitodiplosis mosellana in wheat midge [
3],
Blumeria sp.,
Erysiphe Golovinomyces sp. and
Oidium in powdery mildew [
4],
Puccinia sp. in rust [
5],
Phytophthora infestans in late blight [
6], and
Pseudomonas syringae in bacterial speck [
7]. Plants have developed effective mechanisms to recognize and respond to infections with race non-specific resistance (quantitative) and race-specific resistance (qualitative). Race non-specific resistance involves a number of minor genes that are effective against several pathogens [
8]. For example,
NDR1 and three alleles of
rp1,
rp1-D21,
rp1-MD19 and
rp-NC3, have been shown to confer a race non-specific resistance response to rust [
9]. In contrast, race-specific resistance is conferred by a single or a few major genes that operate in a gene-for-gene fashion in which plant
R-genes recognize corresponding pathogen avirulence (
Avr)-gene effectors to trigger plant defense responses [
10].
Pto in tomato confers race-specific resistance to
Pseudomonas syringae pv. tomato (Pst), similarly to
RPS2 in
Arabidopsis and
N (mosaic virus resistance gene) in tobacco [
11]. On the other hand, the wheat stem rust resistance gene
Sr26 shows resistance to all races of the pathogen
Puccinia graminis f. sp.
tritici, thus constituting a race non-specific pathosystem [
12].
In the defense system of plants, direct and indirect interactions are two alternative mechanisms to explain the gene-for-gene model. In the direct interaction, pathogen Avr effectors associate directly with plant
R-genes to trigger signaling. For example, rice
R-gene
Pi-ta was shown to directly interact with
Avr-Pita from
Magnaporthe grisea [
13]. Likewise, a direct interaction was observed between
L genes (a group of resistant genes to flax rust) and their corresponding rust
Avr genes in flax [
14]. The indirect model is also known as the guard hypothesis [
10]. In this model,
R-gene products act as a “guardee” to monitor the modification of host proteins after associating with the pathogenic effectors, resulting in the initiation of resistance [
15]. Examples of guardee proteins are RPM1 interacting 4 (RIN4), flagellin-sensitive 2 (FLS2), Xa21, resistance to pseudomonas syringae 5 (RPS5) and avrPphB susceptible 1 (PBS1). Resistance gene analogs (RGAs) are a large class of potential
R-genes that have conserved domains and structural features. As such, RGAs can be identified from sequenced genomes using bioinformatics approaches [
16,
17,
18]. In the last 15 years, more than 50 plant genomes have been sequenced and assembled [
19,
20,
21]. Despite this great resource, only a relatively small number of
R-genes have been cloned and fully characterized (
Table 1) providing not only information on their structure, function and evolution, but also generating useful genetic resources to create novel resistant cultivars [
22]. In comparison, thousands of RGAs have been identified in many plant genomes (
Table 2).
Table 1.
Cloned R-genes from plant species.
Table 1.
Cloned R-genes from plant species.
| Species a | R-Gene | Accession ID | Domain b | Class | Chr | Disease c | Avr Gene | Pathogen c | Reference |
|---|
| T. aestivum (wheat) | Lr10 | AAQ01784 | CNL | NBS | 1A | Leaf rust | AvrLr10 | P. triticina | [23] |
| Lr1 | ABS29034 | CNL | NBS | 5D | Leaf rust | Avr1 | P. triticina | [24] |
| Pm3b | AAQ96158 | CNL | NBS | 1A | Powdery mildew | AvrPm3b | B. graminis | [25] |
| Sr33 | AGQ17390 | CNL | NBS | 1D | Stem rust | | P. graminis | [26] |
| Sr35 | AGP75918 | CNL | NBS | 3A | Stem rust | | P. graminis | [27] |
| Lr21 | AAP74647 | NL | NBS | 1D | Leaf rust | AvrLr21 | P. triticina | [28] |
| Cre3 | AAC05834 | NL | NBS | 2D | Cereal cyst | | H. avenae | [29] |
| Cre1 | AAM94164 | NL | NBS | 2B | Cereal cyst | | H. avenae | [30] |
| Yr10 | AAG42168 | CNL | NBS | 1B | Stripe rust | | P. striiformis | [31] |
| Stpk-V (Pm21) | AEF30547 | STK | Oth-R | 6V | Powdery mildew | | B. graminis | [32] |
| Lr34 | ACN41354 | ABC transporter | Oth-R | 7D | Leaf rust, Powdery mildew, Stripe rust | | P. triticina; P. striiformis; B. graminis | [33] |
| Yr36 | ACF33195 | Kinase-START | Oth-R | 6B | Stripe rust | | P. striiformis | [34] |
| H. vulgare (barley) | Mla6 | CAC29241 | CNL | NBS | 1 | Powdery mildew | AvrMla6 | B. graminis | [35] |
| Mla1 | AAG37356 | CNL | NBS | 1 | Powdery mildew | AvrMla1 | B. graminis | [36] |
| Mla13 | AAO16014 | CNL | NBS | 1 | Powdery mildew | AvrMla13 | B. graminis | [37] |
| Rpg1 | AAM81980 | LRR-PK | RLK | 7 | Stem rust | Avr-Rpg1 | P. graminis | [38] |
| Mlo | CAB06083 | TM | Oth-R | 4 | Powdery mildew | | E. graminis | [39] |
| S. lycopersicum (tomato) | Prf | AAC49408 | CNL | NBS | 5 | Bacterial speck | AvrPto | P. syringae | [40] |
| Mi | AAC67238 | CNL | NBS | 6 | Root knot | | M. javanica | [41] |
| I2 | AAB63274 | NL | NBS | 11 | Fusarium wilt | Avr1 | F. oxysporum | [42] |
| Ph-3 | AIB02970 | CNL | NBS | 9 | Late blight | | P. infestans | [43] |
| Sw-5 | AAG31013 | CNL | NBS | 9 | Tomato spotted wilt | | TSWV | [44] |
| Tm-2 | AAQ10735 | CNL | NBS | 9 | Tobacco mosaic | | TMV | [45] |
| Bs4 | AAR21295 | TNL | NBS | 5 | Bacterial spot | AvrBs4 | X. campestris | [46] |
| Hero | CAD29729 | CNL | NBS | 4 | Potato cyst | | G. rostochiensis | [47] |
| Cf-2 | 2207203A | LRR-TM | RLP | 6 | Leaf mold | Avr2 | C. fulvum | [48] |
| Cf-4 | CAA05268 | LRR-TM | RLP | 1 | Leaf mold | Avr4 | C. fulvum | [49] |
| Cf-5 | AAC78591 | LRR-TM | RLP | 6 | Leaf mold | Avr5 | C. fulvum | [50] |
| Cf-9 | CAA05274 | LRR-TM | RLP | 1 | Leaf mold | Avr9 | C. fulvum | [51] |
| Ve1,2 | AAK58681.2 | LRR-TM | RLP | 9 | Verticillium wilt | | V. dahliae | [52] |
| Hcr9-4E | CAA05269 | LRR-TM | RLP | 1 | Leaf mold | Avr4E | C. fulvum | [49] |
| Fen | 2115395A | STK | Oth-R | 5 | Bacterial speck | AvrPto | P. syringae | [53] |
| Pto | A49332 | STK | Oth-R | 5 | Bacterial speck | AvrPto | P. syringae | [54] |
| Pti1 | NP_001233803 | STK | Oth-R | 12 | Bacterial speck | | P. syringae | [55] |
| S. tuberosum (potato) | Rx | CAB50786 | CNL | NBS | 12 | PVX | | PVX | [56] |
| RB | Q7XBQ9 | CNL | NBS | 8 | Late blight | Avr1, Ipio, Ipib | P.infestans | [57] |
| Rx2 | CAB56299 | LZ-NL | NBS | 5 | PVX | | PVX | [58] |
| R1 | AAL39063 | LZ-NL | NBS | 5 | Late blight | Avr1 | P. infestans | [59] |
| L. sativa (lettuce) | Rgc2 (Dm3) | Q9ZSD1 | NL | NBS | | Downy mildew | Avr3 | B. lactucae | [60] |
| P. nigrum (black pepper) | Bs2 | AAF09256 | CNL | NBS | | Bacterial spot | AvrBs2 | X. campestris | [61] |
| O. sativa (rice) | Xa1 | BAA25068 | NL | NBS | 4 | Bacterial blight | AvrXoo | X. oryzae | [62] |
| Pib | BAA76282 | CNL | NBS | 2 | Blast | | M. grisea | [63] |
| Pi-ta | AAK00132 | NL | NBS | 12 | Blast | Avr-Pita | M. grisea | [64] |
| Pi36 | ABI64281 | CNL | NBS | 8 | Blast | Avr-Pi36 | M. grisea | [65] |
| Pia | BAK39926 | CNL | NBS | 11 | Blast | AvrPia | M. oryzae | [66] |
| Pi-Kh | AAY33493 | NL | NBS | 11 | Blast | Avr-Pik | M. oryzae | [67] |
| Pi37 | ABI94578 | NL | NBS | 1 | Blast | | M. grisea | [68] |
| Xa5 | A2XZI2 | NL | NBS | 5 | Bacterial blight | AvrXa5 | X. oryzae | [69] |
| Xa13 | ABD78944 | SET | Oth-R | 8 | Bacterial blight | AvrXa13 | X. oryzae | [70] |
| Pi54 | CCD33085 | CNL | NBS | | Blast | AvrPi54 | M. oryzae | [71] |
| Pi9 | ABB88855 | CNL | NBS | 6 | Blast | AvrPi9 | M. grisea | [72] |
| Piz-t/Pi2 | ABC73398 | CNL | NBS | 6 | Blast | AvrPiz-t | M. grisea | [73] |
| Rpr1 | BAA75812 | CNL | NBS | 11 | Blast | | M. grisea | [74] |
| Pid3 | ACN62386 | CNL | NBS | 6 | Blast | | M. grisea | [75] |
| Xa21 | AAC49123 | LRR-STK | RLK | 11 | Bacterial blight | AvrXa21 | X. oryzae | [76] |
| Xa3/Xa26 | ABD36512 | LRR-STK | RLK | 11 | Bacterial blight | | X. oryzae | [77] |
| CEBiP | BAE95828 | | RLK | 3 | | | | [78] |
| Xa10 | AGE45112 | | Oth-R | 11 | Bacterial blight | AvrXa10 | X. oryzae | [79] |
| Xa25 | AGS56390 | TM | Oth-R | 12 | Bacterial blight | | X. oryzae | [80] |
| Xa27 | AEW90324 | LRR-TM | RLP | 6 | Bacterial blight | AvrXa27 | X. oryzae | [81] |
| Pi-d2 | ACR15163 | B-lectin, STK | RLK | 6 | Blast | | M. grisea | [82] |
| Z. mays (maize) | Rp1-D | AAD47197 | NL | NBS | 10 | Rust | | P. sorghi | [83] |
| Hm1 | Q41867 | | | 1 | Corn leaf blight | | C. carbonum | [84] |
| A. thaliana (Arabidopsis) | RPM1 | CAA61131 | CNL | NBS | 3 | Downy mildew | AvrB, AvrRpm1 | P. syringae | [85] |
| RPS2 | AAA21874 | NL | NBS | 4 | Downy mildew | AvrRpt2 | P. syringae | [86] |
| RPP8/HRT | AAC83165 | CNL | NBS | 5 | Downy mildew | AvrRPP8 | P. parasitica | [87] |
| RPP13 | AAF42832 | CNL | NBS | 3 | Downy mildew | ATR13 | P. parasitica | [88] |
| RCY1 | BAC67706 | CNL | NBS | 5 | Mosaic type | | CMV | [89] |
| RPP1 | AAC72977 | TNL | NBS | 3 | Downy mildew | ATR1 | P. parasitica | [90] |
| RPP4 | AAM18462 | TNL | NBS | 4 | Downy mildew | | P. parasitica | [91] |
| RPS4 | CAB50708 | TNL | NBS | 5 | Powdery mildew | AvrRps4 | P. syringae | [92] |
| RPP5 | AAF08790 | TNL | NBS | 4 | Downy mildew | AvrRp5 | P. parasitica | [93] |
| RPS5 | AAC26126 | NL | NBS | 1 | Downy mildew | AvrRphB | P. syringae | [94] |
| RRS1 | ADM88042 | WRKY-TNL | NBS | 5 | Bacterial wilt | AvrRRS1 | R. solanacearum | [95] |
| RPP27 | CAE51864 | LRR-TM | RLP | 1 | Downy mildew | | P. parasitica | [96] |
| RFO1 | AAY86486 | LRR-STK | RLK | 1 | Fusarium wilt | | F. oxysporum | [97] |
| PBS1 | AAG38109 | STK | Oth-R | 5 | | AvrPphB | P. syringae | [98] |
| FLS2 | AED95370 | LRR-STK | RLK | 5 | Powdery mildew | AvrPto, AvrPtoB | P. syringae | [99] |
| BAK1 | AT4G33430 | LRR-STK | RLK | 4 | | AvrPto, AvrPtoB | P. syringae | [100] |
| NDR1 | AAB95208 | TM | Oth-R | 3 | | AvrB, AvrRpt2 | P. syringae; P. parasitica | [101] |
| RPW8 | AAK09267 | RPW8 | Oth-R | 3 | Powdery mildew | | E. cruciferarum | [102] |
| L. usitatissimum (flax) d | L6 | AAA91022 | TNL | NBS | 5 | Rust | AvrL6 | M. lini | [103] |
| L, L1-L11 | AAD25974 | TNL | NBS | 5 | Rust | AvrBs3 | M. lini | [104] |
| M | AAB47618 | TNL | NBS | 8 | Rust | AvrM | M. lini | [105] |
| P
, P1-4 | AAK28806 | TNL | NBS | 14 | Rust | | M. lini | [106] |
| B. vulgaris (sugar beet) | Hs1pro-1 | AAB48305 | LRR-TM | RLP | 1 | Beet cyst | | H. schachtii | [107] |
| N. tabacum (tobacco) | N | AAA50763 | TNL | NBS | | Tobacco mosaic | | TMV | [108] |
Table 2.
Genome-wide identification of RGAs in plant genomes.
Table 2.
Genome-wide identification of RGAs in plant genomes.
| Species a | Genome Size (Mb) b | Total Annotated Genes b | Total RGAs (%) c | NBS Coding Genes d | PPR e | RLK f | RLP g | Other h | Identification Method Used i | Reference |
|---|
| CNL | TNL | CN | NL | TN | N | Total |
|---|
| Dicots | | | | | | | | | | | | | | | | |
| A. thaliana (Arabidopsis) | 125 | 25,498 | 5.27 | 51 | 79 | 8 | 20 | 17 | 26 | 201 | 441 | 600 | 56 | 46 | H, P, B | [109–113] |
| A. lyrata (lyrata) | 207 | 32,670 | 0.56 | 21 | 103 | 17 | 14 | 20 | 10 | 185 | | | | | H, B | [114] |
| P. trichocarpa (black cottonwood) | 485 | 45,555 | 3.18 | 119 | 64 | 19 | 83 | 13 | 46 | 344 | 600 | 379 | | 127 | MEME, CO, Paircoil2, MC | [111,115–117] |
| V. vinifera (grape) | 475 | 30,434 | 3.81 | 203 | 97 | 26 | 12 | 14 | 0 | 352 | 600 | | | 210 | H, B, MEME | [111,118] |
| L. usitatissimum (flax) | 373 | 43,484 | 0.34 | 31 | 57 | 10 | 5 | 22 | 7 | 132 | | | | 16 | MEME/MAST | [119] |
| S. lycopersicum (tomato) | 900 | 34,727 | 0.84 | 118 | 18 | 19 | 43 | 5 | 49 | 252 | | 16 | 13 | 13 | H, B | [120,121] |
| C. papaya (papaya) | 372 | 28,629 | 0.18 | 4 | 6 | | | | 44 | 54 | | | | | TBN, MEME, CW, MC, H | [122] |
| C. sativus (cucumber) | 367 | 26,682 | 0.26 | 25 | 19 | 1 | 17 | 5 | 3 | 70 | | | | | H, CO, ME, CX, SMART, P, B | [123] |
| S. tuberosum (potato) | 844 | 39,031 | 1.47 | 65 | 37 | 24 | 184 | 12 | 113 | 435 | | | | 142 | H, B | [124] |
| M. truncatula (Medicago) | 454 | 62,388 | 1.20 | 152 | 118 | 25 | 0 | 38 | 328 | 661 | | | | 92 | B, H | [111] |
| G. raimondii (cotton) | 880 | 40,976 | 1.19 | 35 | 41 | 18 | 96 | 9 | 31 | 230 | | 60 | 144 | 56 | B, CO, SMART, MC, CW, IPS, ME5, | [125,126] |
| B. rapa, (chinese cabbage) | 485 | 41,174 | 0.60 | 19 | 93 | 15 | 27 | 23 | 29 | 206 | | | | 42 | B, H | [111] |
| B. oleracea (cabbage) | 630 | 45,758 | 0.52 | 6 | 40 | 5 | 24 | 29 | 53 | 157 | | | | 82 | B, H | [111] |
| F. vesca (strawberry) | 240 | 34,809 | 0.27 | | 61 | | 16 | 8 | 1 | 86 | | | | 8 | B, MU, ME, MEME | [127] |
| M. x domestica (apple) | 742 | 57,386 | 1.86 | 218 | 161 | 54 | 276 | 69 | 182 | 960 | | | | 110 | H, B, CW, MEME | [17] |
| L. japonicus (lotus) | 472 | 19,848 | 0.42 | 9 | 8 | 19 | 3 | 16 | 29 | 84 | | | | | BP, CO, P, MEME | [128] |
| T. cacao (cocoa) | 430 | 28,798 | 1.09 | 82 | 8 | 46 | 104 | 4 | 53 | 297 | | | | 17 | B, H | [111] |
| P. patens (moss) | 510 | 35,938 | 0.46 | 9 | 3 | 2 | 5 | 0 | 1 | 20 | 103 | | | 45 | B, CO, MU, ME | [129,130] |
| Average | 500 | 37,433 | 1 | 69 | 56 | 19 | 55 | 18 | 56 | 263 | 436 | 264 | 71 | 72 | | |
| Monocots | | | | | | | | | | | | | | | | |
| O. sativa (rice) | 420 | 59,855 | 4.22 | 159 | 0 | 7 | 40 | 3 | 45 | 254 | 477 | 1429 | 90 | 281 | H, B, MEME, P | [110,130–133] |
| T. aestivum (wheat) | 17,000 | 94,000 | 2.37 | 98 | | 0 | 555 | | 318 | 971 | | | | 1266 | H, B, MEME | [134] |
| Z. mayes (maize) | 2300 | 32,540 | 0.90 | 58 | 0 | 21 | 31 | 0 | 69 | 179 | | 113 | | 2 | P, H, B, CO | [135,136] |
| S. bicolor (sorghum) | 739 | 34,496 | 1.29 | 36 | 0 | 99 | 133 | 0 | 64 | 332 | | | | 114 | P, H, B, CO, ME, CW | [137,138] |
| H. vulgare (barley) | 5100 | 30,400 | 1.38 | 101 | | 51 | 145 | | 34 | 331 | | | | 89 | | [139,140] |
| B. distachyon (Brachypodium) | 272 | 25,532 | 1.23 | 133 | 0 | 28 | 87 | 0 | 34 | 282 | | | | 34 | P, H, B, CO, CW | [140,141] |
| T. urartu (Red wild einkorn) | 4940 | 34,879 | 1.63 | 235 | 0 | 44 | 218 | | 38 | 535 | | | | 35 | H | [140,142] |
| A. tauschii (Tausch’s goatgrass) | 4360 | 43,150 | 1.94 | 296 | 0 | 63 | 288 | | 81 | 728 | | | | 112 | H | [140,143] |
| Average | 4391 | 44,357 | 2 | 140 | 0 | 39 | 187 | 1 | 85 | 452 | 477 | 771 | 90 | 242 | | |
Though a large number of resistance gene loci have been identified in plants using linkage map** or association studies, most of them correspond to flanking molecular markers or quantitative trait loci (QTL). Mapped genome-wide RGAs, as R-gene candidates, are valuable genomic resources to develop high-density R-gene genetic maps, design diagnostic markers and co-localize QTL. The markers designed from RGAs can be used for fine map** and cloning of R-genes and, for breeding purposes. This review focuses on recent advances in studies of the structures and functions of RGAs, their identification using bioinformatics tools and their applications in genetic research and breeding for disease resistance.
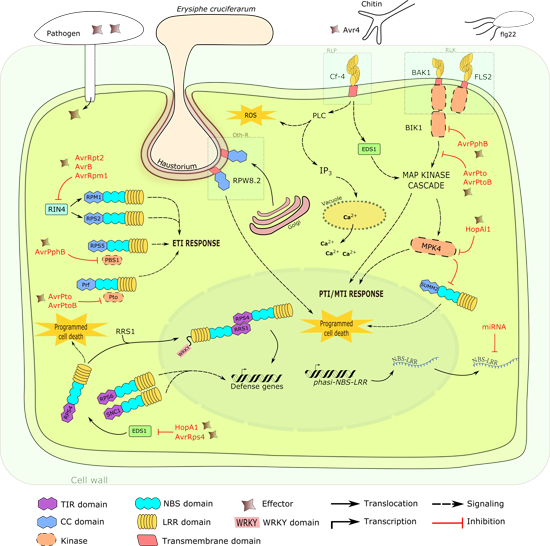
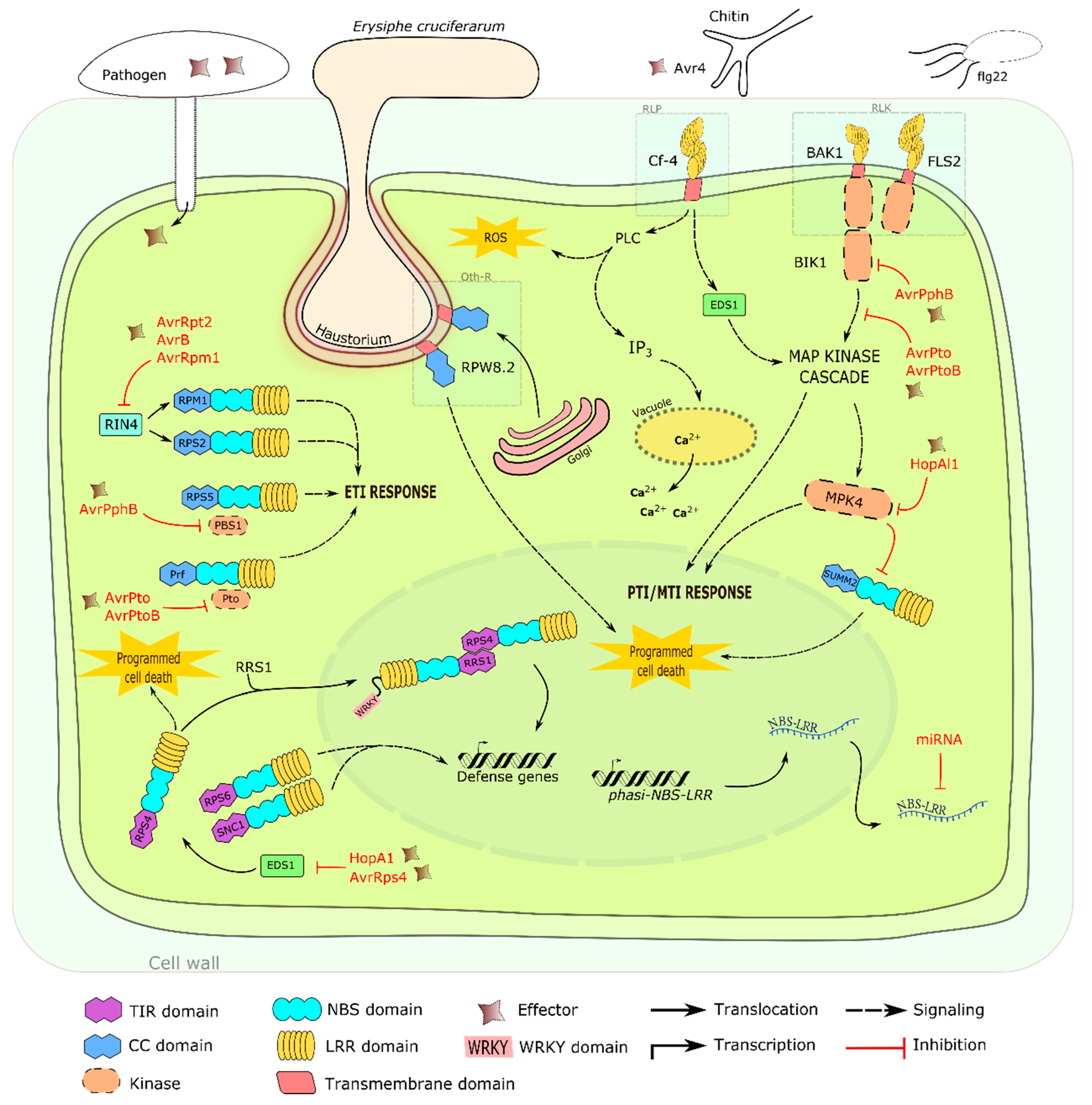
3. Other Defense Related Mechanisms
Aside from the typical NBS-LRR and PPR proteins involved in plant defense, recent discoveries led to the description of other modes of defense. Nomura
et al. [
207] demonstrated that chloroplasts are an important component in activation and signaling of immunity. Upon exposure to flg22, chloroplast calcium-sensing receptor (CAS) dependent immune resistance and PCD are initiated [
207]. Likewise, mitochondria play a similar role [
208], highlighting the importance of these organelles in the defense pathways activated in response to pathogen attacks.
Chloroplast and mitochondrial transmigrated proteins have been described [
207,
208]. They belong to the PPR protein class that constitutes one of the largest protein families in plants. Intriguingly, PPR proteins have undergone an evolutionary process similar with the above-mentioned LRR associated proteins [
209]. Characteristics such as gene clustering and duplications within clusters have been observed [
209]. PPR proteins are nucleus-encoded and will translocate to the chloroplast and mitochondria to perform post transcript processing such as RNA editing, splicing and translation modification [
210].
Known PPR genes, such as
RPF2 and RPF3 in
Arabidopsis,
Rf1 in rice and
Rf2 in maize have been identified ([
211,
212,
213,
214];
Table 3). In addition, an increase in
PPR1 (At1g06580) and
PPR2 (At1g62720) transcripts was observed in response to pathogen attack [
215]. Although their roles were not clearly illustrated, these proteins were speculated to be involved in mitochondrial ROS metabolism [
215]. Likewise, Garcia-Andrade
et al. [
216] identified a negative regulation of PPR protein chlororespiratory reduction21 (CRR21), PPRα (at4g21190) and overexpressor of cationic peroxidase3 (OCP3) in response to chitosan, a fungal PAMP/MAMP. These proteins correspond to chloroplast
NADH dehydrogenase-like complex (
NDH) transcript editing. NDH-mediated immune response remains inconclusive; however, ROS production and callose deposition were speculated to contribute to pathogen-mediated resistance [
216].
Table 3.
Other cloned genes relevant to plant defense.
Table 3.
Other cloned genes relevant to plant defense.
| Species | R-Gene | Accession ID | Domain a | Chr | Disease | Avr | Pathogen b | Reference |
|---|
| H. vulgare (barley) | Rar1 | AAF18432 | CHORD | 2 | Powdery mildew | | B. graminis | [217] |
| S. lycopersicum (tomato) | Asc | AAF67518 | TLC | 3 | Black mold rot; Black shoulder | | A. alternate | [218] |
| O. sativa (rice) | Rf1 | BAC77666 | PPR | 10 | | | | [214] |
| LYP4/6 | | TM | | | | X. oryzae; M. oryzae | [219] |
| Z. mays (maize) | Rf2 | AAC49371 | PPR | 9 | | | | [213] |
| A. thaliana (Arabidopsis) | RPF2 | NP_176454 | PPR | 1 | | | | [211] |
| RPF3 | NP_176481 | PPR | 1 | | | | [212] |
| Rtm1 | AT1G05760 | Jacalin like | 1 | Tobacco etch | | TEV | [220] |
| Rwm1 | AEE33357 | PGK | 1 | Mosaic type | | WMV | [221] |
| EDS1 | AAD20950 | Lipase-like | 3 | | AvrRps4 | P. syringae | [222] |
| NPR1 | AAC49611 | Ankyrin | 1 | | | P. syringae | [223] |
As mentioned above, chloroplast and mitochondria, involved in the production of ROS, activate defense and constitutively initiate PCD [
224]. Various levels of ROS are produced at different concentrations in response to diverse pathogens [
224]. It is intriguing to note that aside from these organelles and oxidase enzymes, about half of the ROS level is generated exclusively from peroxidases upon PAMP/MAMP recognition [
225]. Apoplastic peroxidases, PRX33 and PRX34, have been well studied in response to flg22 and EF-Tu [
225,
226,
227]. Aside from generating hydrogen peroxide (H
2O
2), they play roles in callose deposition and MAMP/PAMP-mediated transcription of defense genes as well [
225,
226]. Therefore, peroxidases are important for plant immunity.
In addition, small RNAs have been found to play a major role in defense, especially in regulating immune components in the cell [
174]; however, further studies are needed as their mechanisms remain poorly characterized.
4. Bioinformatics Approaches for RGA Identification and Characterization
To date, genome sequences of more than 50 plant species have been sequenced and assembled to various degrees [
19,
20,
228]; the released sequences were deposited in public databases such as Phytozome [
19,
20] and EnsemblPlants FTP servers [
229]. Advances in next generation sequencing technologies have made whole genome sequencing one of the most important approaches in modern biological research. Current challenges include the provision of functional annotations for the large number of macromolecules. However, experimental investigations to assign protein functions are costly and time consuming. Alternatively, computational approaches to functional prediction are very attractive to solve this complex task [
230]. Mining and characterizing genome-wide plant RGAs using computational approaches are rendered possible due to their significant structural features and conserved domains. Several bioinformatics methods have been applied to identifying RGAs and predicting their functions, including sequence alignment, BLAST search, phylogenetic analysis, and domain and motif analysis [
231] using several applications such as Hidden Markov Model (HMM) [
232], SMART (
http://smart.embl-heidelberg.de/) [
233], Prosite (
http://prosite.expasy.org/), pfam (
http://pfam.xfam.org/), and InterProScan5 (
http://www.ebi.ac.uk/Tools/pfa/iprscan5/) which are summarized (
Table 2 and
Table 4).
Based on previously used approaches, the identification and characterization of RGAs usually follow a common procedure of four steps (
Figure 3). First, a plant RGA database including all known plant RGA gene and protein sequences is generated. GenBank [
234] and PRGdb [
235] are two important sources of well curated RGA sequences. Second, BLAST searches against the RGA database are performed using a loose E-value cut-off (from 1e-5 to 1 depending on the genome size) to identify RGA candidates. Third, using the RGA candidates as input, a variety of software tools (
Table 4) are employed to detect various conserved domains and motifs and produce alignments. Some programs like pfam_scan.pl (developed by Sanger) and InterproScan can be run in a parallel mode. In the last step, a dedicated sorting script is needed to group the RGA candidates into classes as per their domain and motif structures or a combination thereof. For example, to be classified as a gene encoding a TNL protein, an RGA must have a 5′ TIR and an NB-ARC followed by an LRR domain.
To date, no standardized bioinformatics tools and consistent annotation criteria were employed in individual studies. Also, individual software tools may have their own advantages and limitations in identifying different types of RGA domains. Thus, the results from different studies are not necessarily comparable. A comprehensive pipeline package to seamlessly integrate these individual tools is expected to save biologists’ time by facilitating processing, standardizing data organization and providing visualization features. The use of consistent criteria to identify the complete RGA complements would permit their comparative analyses across species.
Table 4.
Common software used for RGA domain and motif identification.
Table 4.
Common software used for RGA domain and motif identification.
| Software | Latest Version | Input Type a | Required Database | Description | Parallel Support b | URL c | Reference |
|---|
| HMMER | 3.1b2 | D/P | HMM model | Protein or DNA sequence homolog search toolkits using profile hidden Markov models and featured by remote homolog identification. The latest version is as fast as BLAST thanks to the underlying mathematical models. | HT/MPI | hmmer.janelia.org | [236] |
| MEME | 4.10 | D/P | | Discover novel and ungapped motifs from nucleotide or protein sequences without well trained dataset samples. | MPI | meme-suite.org | [237] |
| mCUDA-MEME | 3.0.15 | D/P | | An ultrafast scalable motif discovery program running on graphics processing unit (GPU). The algorithm is based on MEME using a hybrid combination of CUDA, MPI and OpenMPI parallel programming models. | CUDA/MPI | bit.ly/18X8LmA | [238] |
| BLAST+ | 2.30 | D/P | BLAST databases, like nr or nt database | Classical similarity search toolkits for bioinformatics data mining. The latest version significantly improves the speed on CPU and efficiency on RAM for long queries. | HT | blast.ncbi.nlm.nih.gov | [239] |
| pfam_scan.pl | 1.0 | P | Pfam-A HMM model | A Perl script for PFAM database search, which invokes “hmmscan” in the HMMER toolkit package to search known HMM models. | | bit.ly/1M41KRu | |
| InterproScan | 5.9 | P | PFAM, SMART, PANTHER, PROSITE, Superfamily,
etc. | A tool that combines different protein signature recognition methods native to the InterPro member databases into one resource with lookup of corresponding InterPro and GO annotations. | HT | www.ebi.ac.uk/interpro | [240] |
| Phobius | 1.01 | P | HMM model | A HMM based tool for transmembrane (TM) topology and signal peptides (SP) prediction from proteins. A pre-training HMM model has been embedded in the tool. | | phobius.sbc.su.se | [241] |
| TMHMM | 2.0 | P | HMM model | A HMM based tool with similar functions to Phobius. | | www.cbs.dtu.dk/services | [242] |
| nCOILS | 2.2 | P | Scoring matrix | A program to detect CC domains by comparing and scoring protein sequences with a known coiled-coils database with the MTK or MTIDK calculation matrix, which reports a probability that the sequence adopts a coiled-coil conformation. | | embnet.vital-it.ch | [243] |
Figure 3.
A common procedure for identification and characterization of plant RGAs.
Figure 3.
A common procedure for identification and characterization of plant RGAs.
5. Genome-Wide Identification and Characterization of RGAs
Whole genome sequencing of plants has enabled genome-wide identification, map** and characterization of RGAs. NBS-LRR containing RGAs have been evaluated in numerous plants such as apple,
Arabidopsis, barley, black cottonwood, grape, maize,
Medicago, rice and sorghum (
Table 2). Hundreds of NBS-LRR encoding genes have been identified from these plant genomes. NBS-LRR genes are a highly duplicated, evolutionarily diverse and clustered gene family [
244] and, they represent the major class of
R-genes contributing to disease resistance in plants [
245]. Angiosperms possess NBS-LRR encoding genes but TNL encoded genes are absent from grass genomes [
196,
246] and other monocots [
114]. It has been hypothesized that this absence is likely due to either a loss or the lack of amplification of TNL encoding genes in the monocot lineage [
156,
247,
248]. The loss of TNL-encoding genes in monocots could be due to increased dependence on CNL proteins [
156]. In rice, CNL proteins are encoded by many more genes than the total number of CNL and TNL genes in
Arabidopsis [
156]. More NBS-LRR and CNL encoding genes have been identified in most monocots than in dicots (
Table 2). CNL and TNL proteins may utilize different downstream signaling components in disease resistance responses [
249]. This genomic shift to CNL genes in monocots could have resulted from mutations in genes encoding downstream components of TNL-specific pathways, resulting in a loss of function and conservative selection for TNL genes. Therefore, TNL-encoding genes may have been lost or simply never amplified in monocot genomes due to a lack of selective advantage [
156]. In addition, some dicots like
Arabidopsis contain more TNL than CNL [
109,
118]. The abundance of TNL likely results from its older origin than CNL [
250]. Besides TNL and CNL, variants of NBS encoding genes such as CN, NL, TN and N vary largely in number (
Table 2). Other NBS-LRR like domains such as CTNL and CTN in apple [
17], TN-TNL, TTNL, XTNX and SA-CA (signalling commentator with CC domain, e.g., RPW8) in
Arabidopsis [
251], and, CNLX, CNX, CNXL, CXN, NX and NLX in sorghum [
138], were reported. Numbers of TIR-X RGAs were also reported such as 126 in cabbages, 46 in
Arabidopsis, 67 in cottonwood and 92 in
Medicago [
111].
More than 600 members of RLKs were identified in
Arabidopsis [
112] and around 1200 members in rice [
132]. They have also been reported in maize, wheat, tomato and cottonwood (
Table 2). RLPs with TM domain have also been reported in
Arabidopsis and tomato (
Table 2). Some cloned genes, such as
Hs1pro-1 in sugar beet,
Cf2,
Cf4-9,
Ve1 and
Hcr9-4E in tomato, belong to this class (
Table 1).
Some RGAs have been identified as pseudogenes. A total of 49
R-pseudogenes in
Medicago [
16], 179 in potato [
124], 347 in the rice variety “Nipponbare” and 345 in variety “93-11” [
252], 10 in
Arabidopsis [
109], 161 in cottonwood [
115] and 62 in lotus [
128] have been identified. In tomato, only 10% of the cloned RGAs were classified into pseudogenes [
253], but almost half of the identified RGAs were pseudogenes in western white pine (
Pinus monticola) [
254]. Pseudogene paralogs of several R-genes such as
Xa21,
Cf9,
Pto and
Dm3, were also identified [
254]. Most identified pseudogenes have strong identity with another NBS protein but their sequences are shortened by premature stop codons or frameshift mutations. Sequence comparisons have suggested that RGA pseudogenes originated from point mutations, like insertion or deletion of nucleotides [
254]. Usually, pseudogenes are considered non-functional genes; however, some pseudogenes can be transcribed into mRNA [
254,
255]. Evidence of expression also exists in species such as rice [
256], pine [
254] and
Medicago [
16]. In these species, some pseudogenes have 90%–100% identity to ESTs and their functions are ascribed as partial NBS-LRR proteins. Pseudogenes are believed to power
R gene evolution by driving illegitimate recombination and permitting gene conversion with alleles or paralogs of functional
R-genes [
257].
As noted earlier, there is surprising similarity in the
PPR genes of
Arabidopsis and rice supporting their existence prior to the divergence of monocots and dicots [
130]. However, massive differences between the numbers of
PPR genes in higher plants and non-plant organisms are indicative of the expansion of this gene family during the evolution of the plant kingdom [
130].
Some RGAs represent a highly divergent defense system in plants, containing a large and diverse family of genes with conserved motifs and domains [
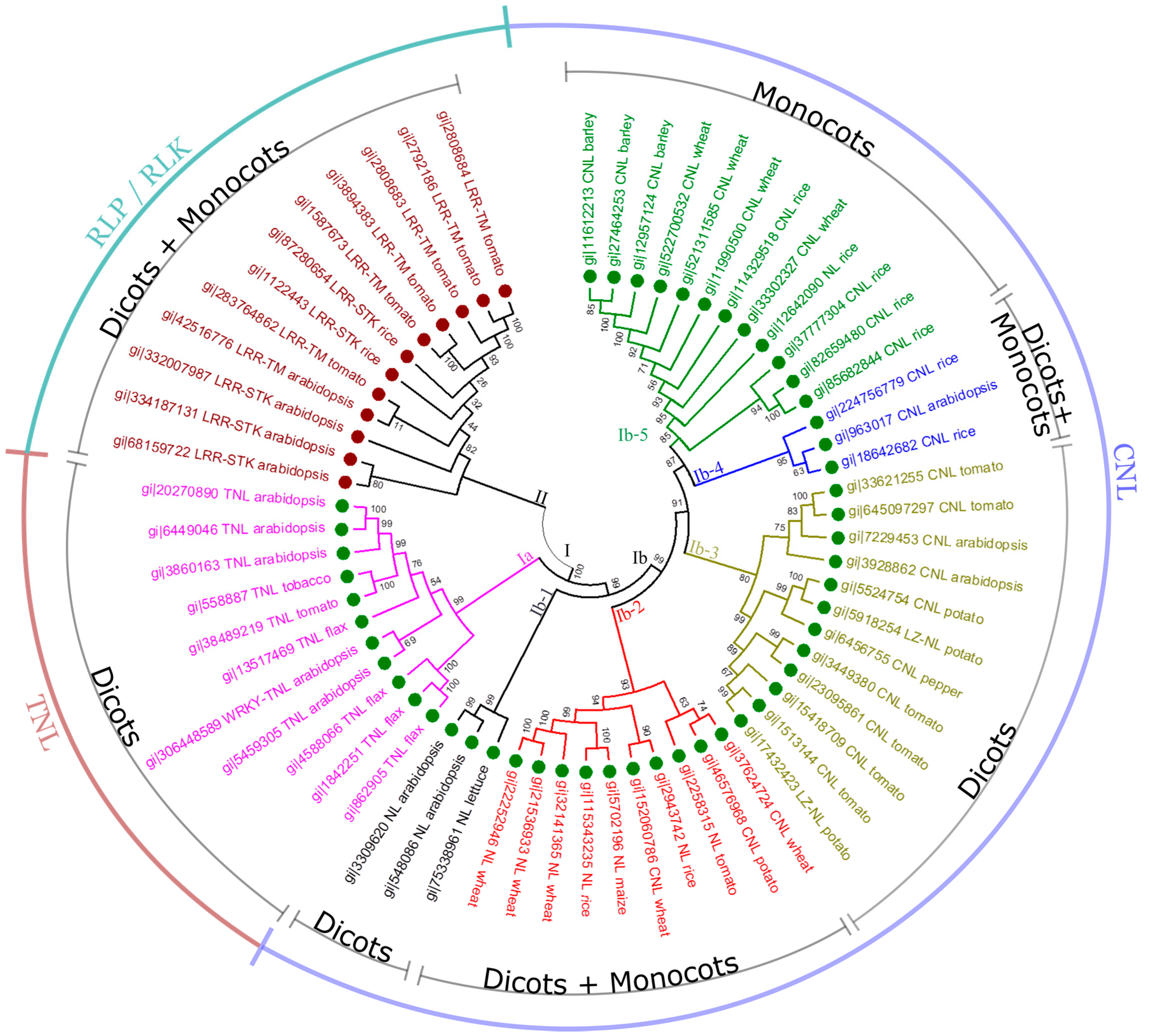
254]. To understand the phylogenetic relationship of RGAs, we selected 63 well-characterized RGAs or cloned
R-genes (
Table 1) in seven dicot (
Arabidopsis, black pepper, flax, lettuce, potato, tobacco and tomato) and four monocot species (barley, maize, rice and wheat) from the NCBI database. These selected RGAs represent the major classes of RGAs: NBS-LRR (CNL, TNL or NL), RLK (LRR-STK) and RLP (LRR-TM). A neighbor-joining tree with 63 RGAs was constructed using MEGA 6 [
258] (
Figure 4). RGAs were classified into two groups: Clade I for NBS-LRR encoding genes and Clade II for RLK and RLP encoding genes. As RLP and RLK proteins have common domains but differ in presence or absence of a kinase (
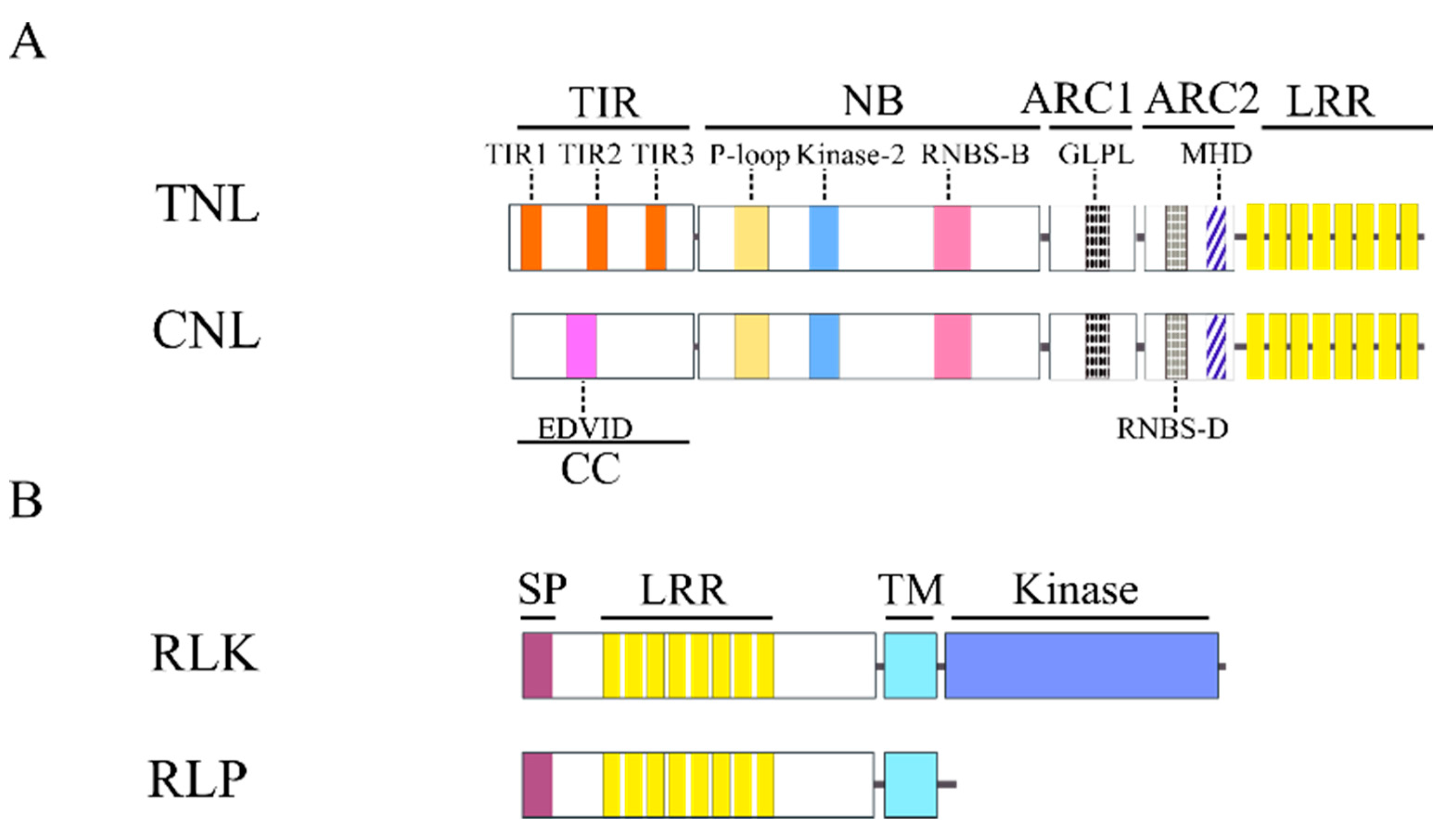
Figure 1B), LRR-STK and LRR-TM encoding genes clustered into one large group (Clade II) with sub-clusters. CNL and TNL are major subclasses of NBS-LRR encoding genes; thus they grouped into distinct sub-clusters in Clade I (Clades Ia for TNL and Ib for CNL encoding genes, respectively) (
Figure 4). We observed that TNL encoding genes were conserved among different dicot species (Clade Ia). In contrast, CNL encoding genes are much more diverse. Some of them are conserved within angiosperms (Clades Ib-2 and Ib-4), within dicots (Clades Ib-1 and Ib-3) or within monocots (Clade Ib-5), while others have diverged between dicot and monocot species (Clade Ib) or within dicots (between Clades Ib-3 and Ib-5). For example, Clade Ib-3 contains RGAs from dicot species only while Clade Ib-5, from monocot species only. In addition, NL and LZ-NL encoding genes may be more closely related to CNL than to TNL because almost all NL and LZ-NL encoding genes clustered with the CNL clades (Clades Ib-1, Ib-2, Ib-3, Ib-4 and Ib-5), suggesting that the variants of non-TNL, such as NL and LZ-NL, may have evolved from CNL rather than TNL.
Figure 4.
Phylogenetic analysis of RGAs in plants. The protein sequences of 63 RGAs or cloned
R-genes from eleven plant species were selected for this analysis using MEGA 6 [
258]. The protein sequences were aligned using the Muscle algorithm, and then clustered using the neighbor-joining algorithm with the p-distance model, pairwise deletion for gaps or missing data treatment, and 500 bootstrap replicates. The phylogenetic tree consists of two large clades (Clades I and II), representing the NBS-LRR class and the RLK/RLP class of proteins, respectively. Clade I may be divided into two sub-groups (Clades Ia and Ib), containing TNL and CNL proteins, respectively, while Clade Ib may be further split into several diverged CNL clusters. The bootstrap values are labelled on branches.
Figure 4.
Phylogenetic analysis of RGAs in plants. The protein sequences of 63 RGAs or cloned
R-genes from eleven plant species were selected for this analysis using MEGA 6 [
258]. The protein sequences were aligned using the Muscle algorithm, and then clustered using the neighbor-joining algorithm with the p-distance model, pairwise deletion for gaps or missing data treatment, and 500 bootstrap replicates. The phylogenetic tree consists of two large clades (Clades I and II), representing the NBS-LRR class and the RLK/RLP class of proteins, respectively. Clade I may be divided into two sub-groups (Clades Ia and Ib), containing TNL and CNL proteins, respectively, while Clade Ib may be further split into several diverged CNL clusters. The bootstrap values are labelled on branches.
6. Genome Organization of RGAs
Many studies revealed that RGAs are irregularly distributed in plant genomes and that many reside in clusters. The clustered distribution of RGAs provides a reservoir of genetic variation to drive the evolution of new
R-gene specificities [
257,
259,
260].
Most RGAs are found in clusters. For example, 38.2% of the mapped NBS genes were located into eight clusters in the lotus genome [
128]. Similarly, 50% and 51% of NBS genes were clustered in the rice [
261] and
Brachypodium genome [
141], respectively. Higher rates have been found in other species such as potato where 73% of the mapped NBS-LRR genes grouped into 63 clusters [
262], while approximately 80% were clustered in
Medicago [
16], 74.3% in the rice cultivar “Nipponbare” and 71.1% and 63.8% in
Arabidopsis and lyrata, respectively [
114]. Many super
-clusters were identified for NBS-LRR genes, including a super
-cluster with 11 NBS genes on chromosome 4 of
Brachypodium [
141] and one with 82 NBS-encoded genes in
Medicago [
16]. In addition, some
R-genes appear in complex cluster structures [
257] forming a diverse multigene family such as
Cf4/9 in tomato [
49],
L in flax [
14],
Mla in barley [
263],
Pi2/9 in rice [
260],
Dm3/13 in lettuce [
60] and
I2 in tomato [
42]. The structures of three haplotypes (linked genes) of the
Pto cluster in tomato have been defined [
264] while a single haplotype of the
Xa21 cluster in rice was partially characterized [
265]. Several restorer genes, like the
Rf gene from various plant species, showed homology to a cluster of
PPR genes [
266]. Genes within a single cluster may determine resistance to different pathogens [
257].
RGAs are often irregularly distributed on chromosomes. Gebhardt
et al. [
267] and Lozano
et al. [
124] published genetic maps of potato with resistance traits. Their analyses indicated that large numbers of NBS-LRR genes were found on chromosomes 4 and 11 (approximately 15% of the mapped genes), while only 1% was on chromosome 3. In
Medicago, chromosomes 6 and 3 encoded approximately 34% and 40% of all TNLs, respectively [
16]. In apple, 56% of the 868 identified RGAs were distributed on six chromosomes (Chr 2, 7, 8, 10, 11 and 15) including 25% on Chr 2 while 99% of the non-TNL class was on Chr 11 [
268]. Pseudogenes are also distributed and clustered at specific chromosomal locations in the same way as functional NBS-LRR genes [
16,
124]. Pseudogenes evolve much faster than functional genes and represent a reservoir for evolution of new specificities [
269].
In conclusion, genetic and molecular data have shown that disease resistance genes are frequently clustered in plant genomes. Several cloned
R-genes have common domains which will help to identify disease resistance loci from clusters of paralogs [
257].
R loci may harbor single genes with multiple alleles, for instance, the
L locus in flax with 13 alleles or
RPM1 in
Arabidopsis with two [
257]. However, some resistance loci and clusters of resistance genes are inherently unstable, e.g.,
Rp1 in maize was described as a fast evolving complex [
270].




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



