1. Introduction
Polyamines (PAs) are low molecular weight aliphatic amines containing two or more amino groups found in all living cells [
1]. In higher plants, PAs are mainly present in their free form, and have several potent biological activities. Thus, PAs are considered to be a new group of growth regulators in plants [
2]. The total and individual PAs levels vary depending on the plant species, the organ and on the developmental stage; furthermore, their metabolism is dynamic, due to the polyamine cycle [
3]. Experiments on treatments with both PAs and PA synthesis inhibitors revealed the role of PAs in e.g., flower induction [
4], embryogenesis [
5], regulation of nucleic acid synthesis and protein translation, development of organelles [
6], and senescence [
7], and numerous studies have been published on the relationship between the enhanced synthesis of PAs and the level of a stress tolerance [
8,
9,
10]. A positive role of the early stress-induced activation or the overexpressing of certain PA-biosynthesis genes has also been proved (arginine decarboxylase:
ADC, spermidine synthase:
SPDS and spermine synthase:
SPMS) [
11,
12,
13,
14,
15,
16]. In addition, microarray studies have shown that increased endogenous PA content can alter the gene expression level of genes involved in the synthesis and signalling of several plant hormones, such as abscisic acid (ABA), auxin, ethylene, gibberellins (GAs), jasmonic acid (JA) or salicylic acid (SA) in transgenic
Arabidopsis plants [
17]. It is thus becoming more evident that PAs are also switching points in plant signalling pathways, and the induced plant responses are interconnected at many levels with other metabolic routes and hormonal cross-talk and activate gene expression, which has a predominant role in the PA-signalling processes compared only to PA accumulation [
18].
Most studies have focused on the protective effects of exogenous PAs, but their role is more complicated. It is difficult to establish a direct relationship between PAs, especially the individual levels of the most abundant Pas—namely putrescine (PUT), spermidine (SPD) and spermine (SPM)—and the observed beneficial effects. In addition, an excess of PAs can be harmful to the plants [
15]. According to these facts, changes in the PA levels are important for fine-tuning PA signalling, which influences the hormonal balance required for them to exert their positive role in regulating plant growth and stress tolerance [
15].
Earlier it was demonstrated that SA treatment affects the synthesis and/or catabolism of PAs [
19,
20,
21,
22]. Vice versa, it was shown that SPD treatment increased SA content in the leaves of wheat, but PUT or SPD treatments decreased it in the roots [
23], whereas SPD did not affect endogenous SA content in cucumber [
24]. In our previous study, it was found that there is a close, positive relationship between PAs and SA accumulation after SPD and SPM treatments in wheat and maize plants [
15]. SA synthesis starts from chorismate as a precursor, after which the synthesis pathway branches into two separate pathways. SA can be synthesized through the phenylalanine ammonia-lyase (PAL) pathway from phenylalanine via benzoic acid (BA), while on the isochorismate pathway the enzyme isochorismate synthsase (ICS) catalyzes isochorismate formation from chorismate. In
Arabidopsis, the primary route for SA production is the isochorismate pathway, and there are two genes
ICS1 and
ICS2 encoding ICS, but it was demonstrated that ICS1 is responsible for the main source of isochorismate [
25]. Excessive SA accumulation can be detrimental to plants under stress conditions. For example, a negative relationship was observed between the SA content and growth parameters of
cpr1 Arabidopsis mutant plants, which showed higher levels of free and bound SA and increased oxidative damage under low temperature stress conditions [
26]. Increased SA accumulation was also responsible for the negative effect of SPD and SPM treatments, especially at higher concentration, manifested in growth inhibition in wheat and maize under control conditions [
15], and also for the accelerating effect of SPD treatment on cadmium-induced oxidative stress in wheat [
16].
However, during the investigation on the relationship between SA and PAs, it should be also taken into consideration that SA may also influence other plant hormones, e.g SA has an antagonistic relation with ABA and JA [
27,
28], thus the PA-induced hormonal changes may also be affected.
A positive feedback loop has also been found between ABA and PAs, as ABA activates the PA synthesis genes and also that of polyamine oxidase (
PAO); moreover PA treatment induces ABA synthesis at the gene expression level of 9-
cis-epoxycarotenoid dioxygenase (
NCED) [
29,
30,
31]. In a recent study, PA treatments differently influenced the plant growth parameters of GA-insensitive dwarf (carrying the severe dwarfing allele
Rht-B1c, responsible for dwarf phenotype) and semi-dwarf Rht (carrying
Rht-B1b allele) wheat lines compared to the wild, tall line, which effects were in relation with different changes in ABA and SA contents of the three genotypes [
32]. These results provide new insights into the role of PAs in plant growth regulation and confirmed their participation in the hormonal balance, however, still little is known about the interplay between PAs and GAs. Methyl jasmonate treatment in barley induced the expression of genes involved in PUT synthesis
ADC and ornithine decarboxylase, which in turn led to increased PA content in the conjugated fraction [
33].
OsPAO6 has been also reported to be induced by JA [
34]. Exogenous PAs, especially SPM, induced JA accumulation after only 1 h of treatment [
35].
In order to reveal even more deeply the relationship between PAs and SA at the metabolite and gene expression levels, and the contribution of SA synthesis deficiency to the interplay of PA with other plant hormones, such as ABA, GAs and JA, in the present study,
eds5 and
sid2 mutants of
Arabidopsis—which do not accumulate SA after pathogen infection or abiotic stresses [
36,
37]—were tested after PUT, SPD or SPM treatments. Our hypothesis was that the different Pas would induce different changes in the PA and SA metabolism of the wild type and SA-deficient
Arabidopsis mutants, which in turn would influence other plant hormones. To our knowledge, such a side-by-side comparison of the influence of
eds5 and
sid2 mutation on the effect of different PAs (PUT, SPD and SPM) has not been reported yet. The main questions were: (1) Is there any difference in the PA metabolism between the SA-deficient mutants and the wild type? (2) How do exogenous PAs influence SA synthesis? and (3) Does SA-deficiency have any influence on the PA-induced changes in the ABA, JA and GA contents?
3. Discussion
The main aims of the present study were to reveal the possible effects of the eds5 and sid2 mutations on the polyamine metabolism in Arabidopsis plants, and to evaluate the effects of exogenous PA on SA synthesis, and the influence of SA-deficiency on the PA-induced hormone synthesis.
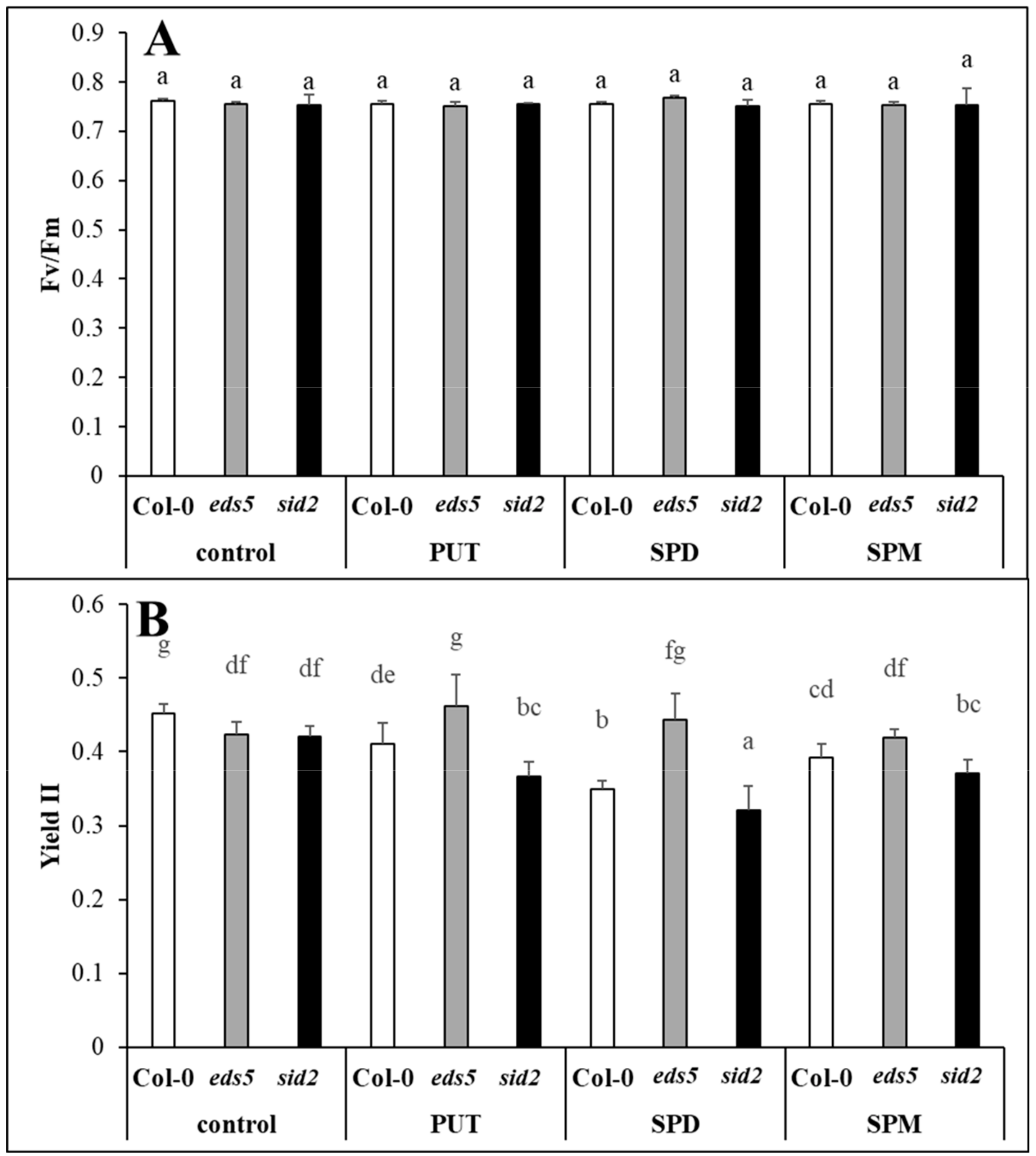
Besides the visually observed phenotypical differences between the two SA-deficient mutants and wild type, the Y(II) parameter also showed that the two mutants have lower PS II quantum efficiency compared to the Col-0. Similarly, although no differences were observed in the Fv/Fm, but slightly lower effective PSII quantum yields were measured for the SA-deficient
sid2 mutant and
NahG transgenic line compared to the wild type [
38]. Parallel with these, slightly lower stomatal conductance and CO
2 assimilation rate, and lower biomass parameters were measured for
sid2 mutant. These results confirmed that controlled levels of SA are required for optimal photosynthesis. However, SA deficient mutants, namely
eds5 and
sid2 have not been tested under PA treatments yet. Under the present conditions, PA treatments repressed the efficiency of PSII, based on the Y(II) parameter, especially in Col-0 and
sid2 genotypes, while a slight positive effect of PUT treatment has been detected in the case of
eds5 mutant compared the its control. PUT treatment at the same concentration has been reported to have a corroborating effect under control conditions in wheat plants, manifested in higher shoot fresh and dry weight and CO
2 assimilation rate, and the beneficial effect of PUT was accompanied with the lowest SA accumulation [
15]. In the present study, PAs induced the lowest decrease in Y(II) in the case of
eds5, where interestingly the lowest SA values were detected.
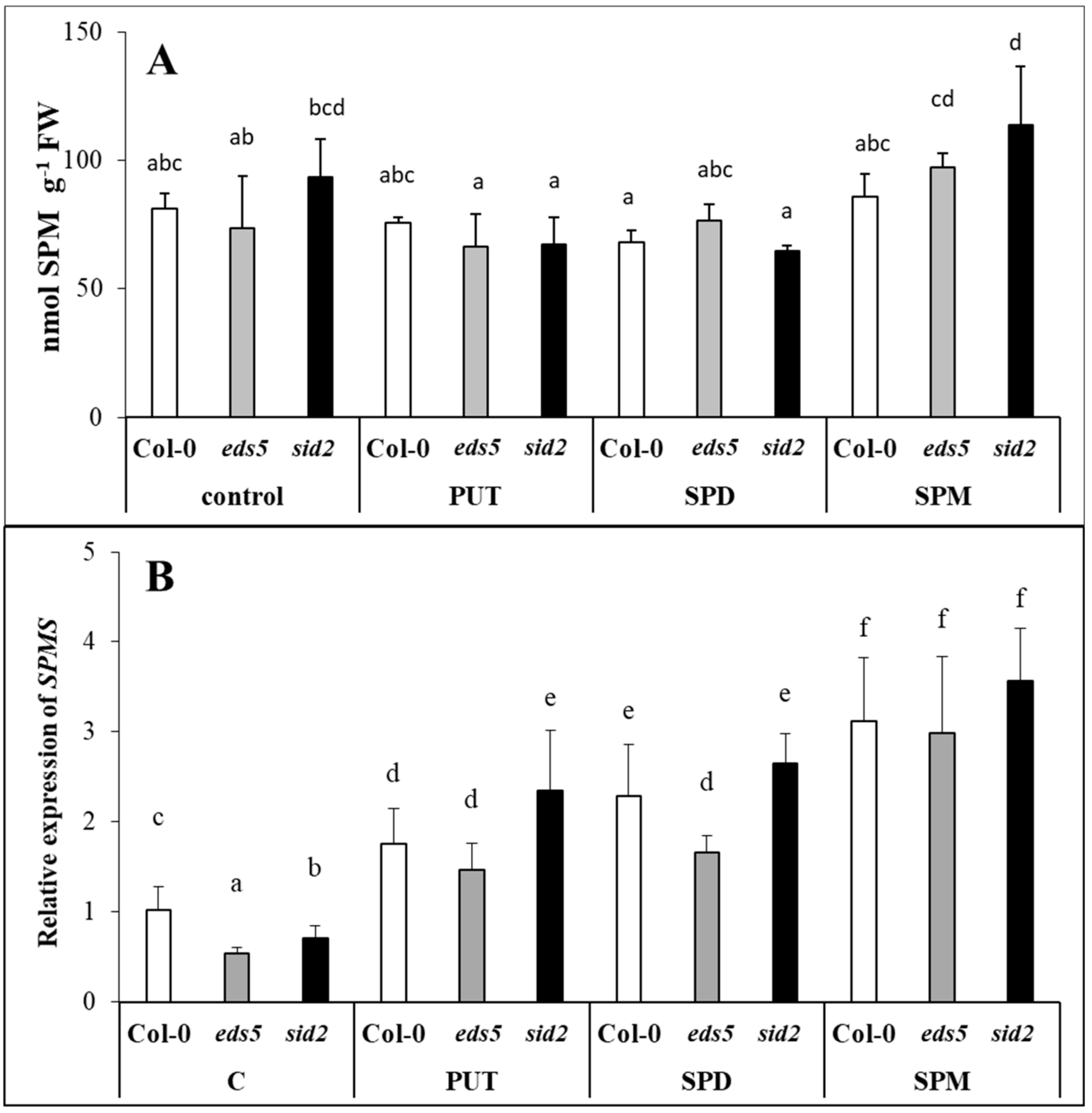
Under control conditions, the
eds5 and
sid2 mutations did not influence the PA contents, among the genes involved in PUT, SPD and SPM synthesis, back-conversion or terminal catabolism, only the expression levels of SPMS showed higher transcript level in
sid2 compared to the
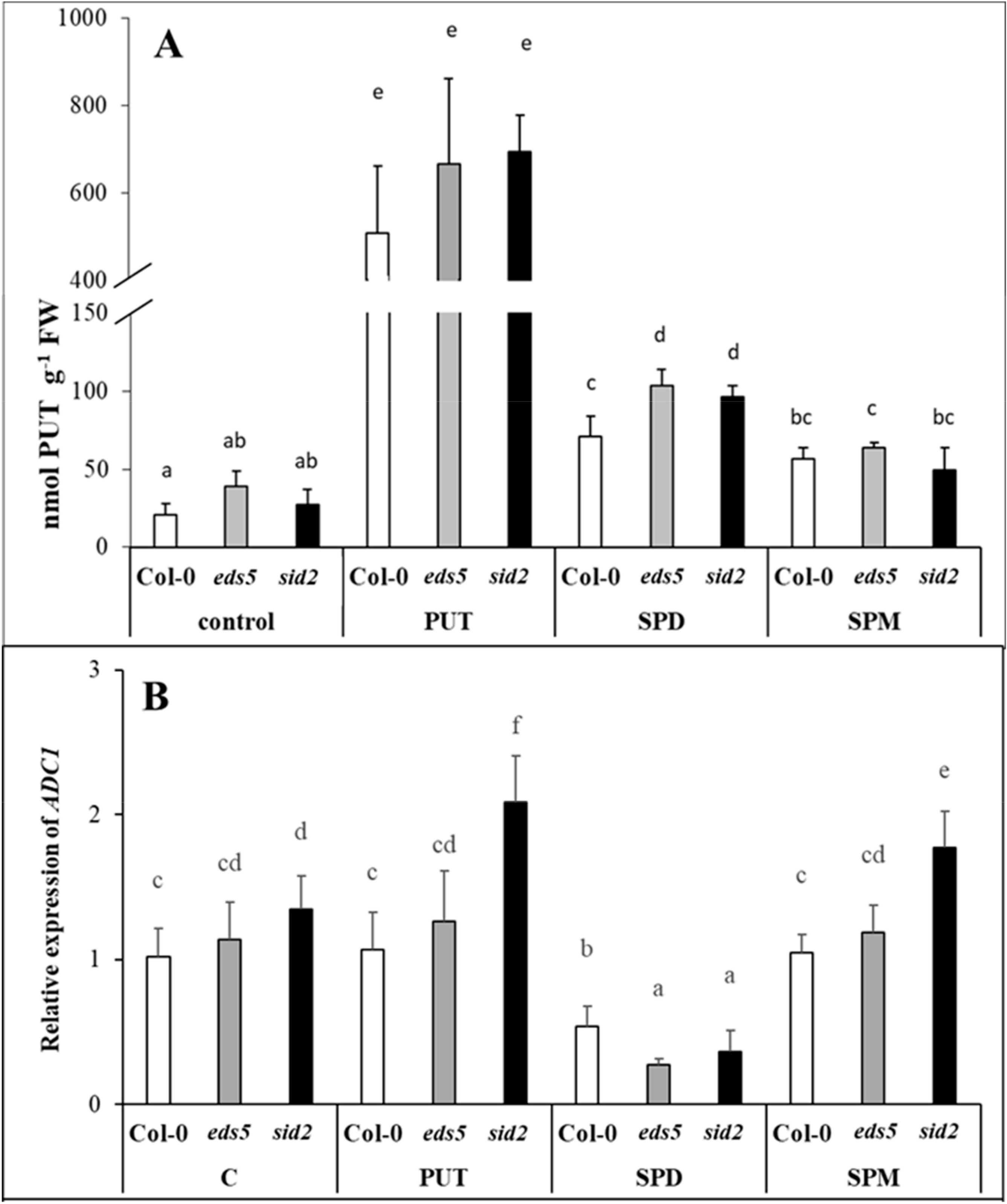
eds5 and the wild type. PA treatments induced remarkable increments only in the PUT content, but these were similar in the three genotypes, with the highest values in the case of PUT-treated plants. Besides the PUT uptake and translocation into the leaves, which was predominant in PUT-treated plants,
de novo PUT synthesis was also occurred, as the expression level of
ADC2 was significantly induced by all the PA treatments, especially after SPD and SPM treatments. Our results regarding the different inducibility of the expression of
ADC1 and
ADC2, are in accordance with the literature, as it seems that
AtADC1 is constitutively expressed, while
AtADC2 is responsive to abiotic stresses, plant hormones or pathogens [
39]. Interestingly, under the present conditions, the initial transcript level of
ADC1 was slightly higher in
sid2, and this difference became more pronounced after PUT and SPM treatment, while
ADC2 expression level was higher in
sid2 mutants after SPD or SPM treatments compared to the wild type. The
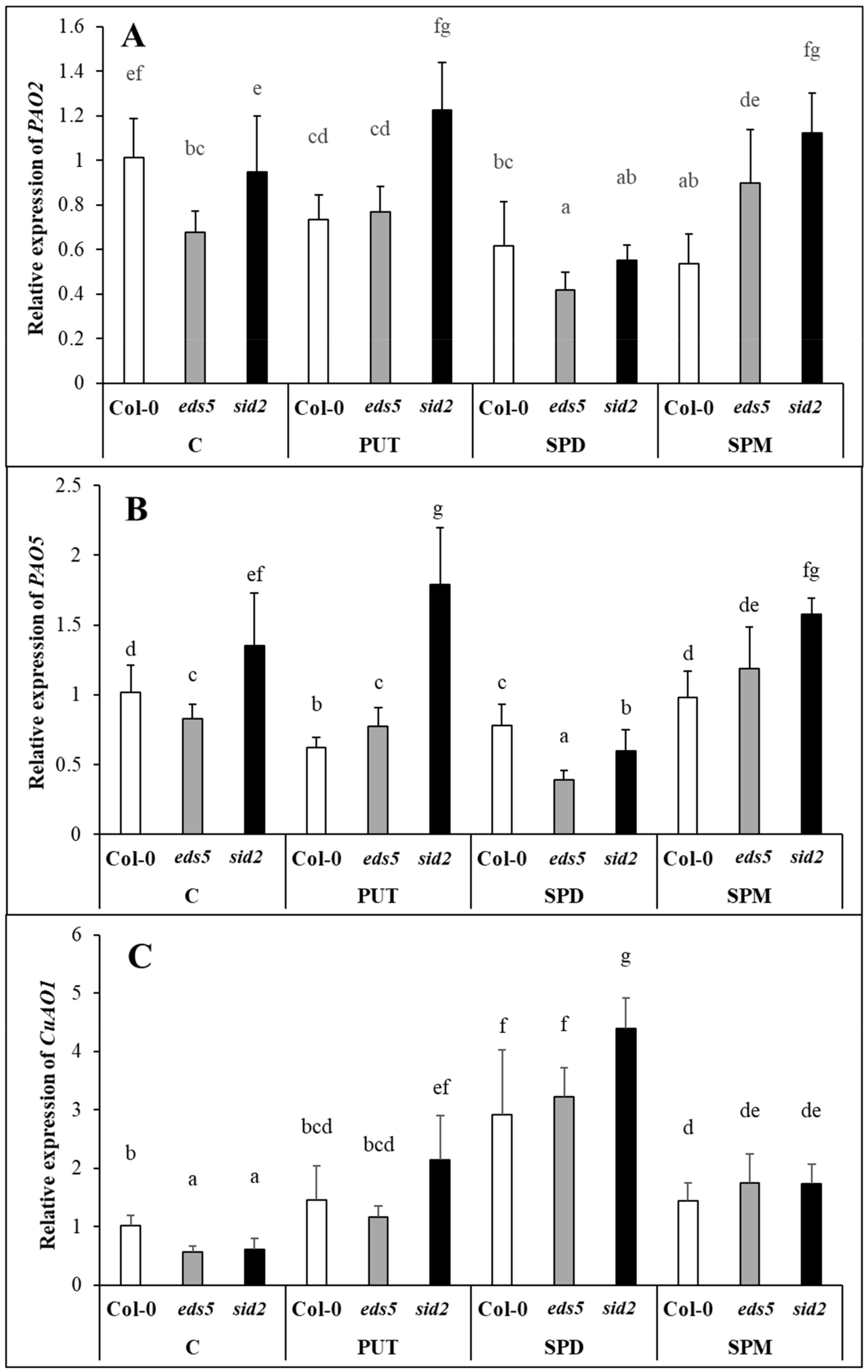
PAO2 and
PAO5 expressions in the present experiment showed exactly the same pattern, and did not show remarkable changes, except for in SPD-treated plants, where they were down-regulated. As
AtPAO2 and
AtPAO5 encoding polyamine oxidases [
40,
41], which are responsible for the back-conversion of higher PAs to PUT, it is not surprising that their expression was not induced upon increased PUT content and there was no need for back-conversion [
42]. Except for SPD treatment, the expression level of the
PAOs, similarly to the genes of PUT synthesis, was the highest in the
sid2 mutant. At the same time, the expression of
CuAO1, that encoding an amine oxidase catalyses the terminal catabolism of PUT and SPD, increased after all the PA treatments, especially in case of SPD, the most pronouncedly in the
sid2 genotypes. Although, the activation of terminal catabolism was not sufficient to compensate the increased PUT content, and the differences in its pattern were not manifested in the PUT contents.
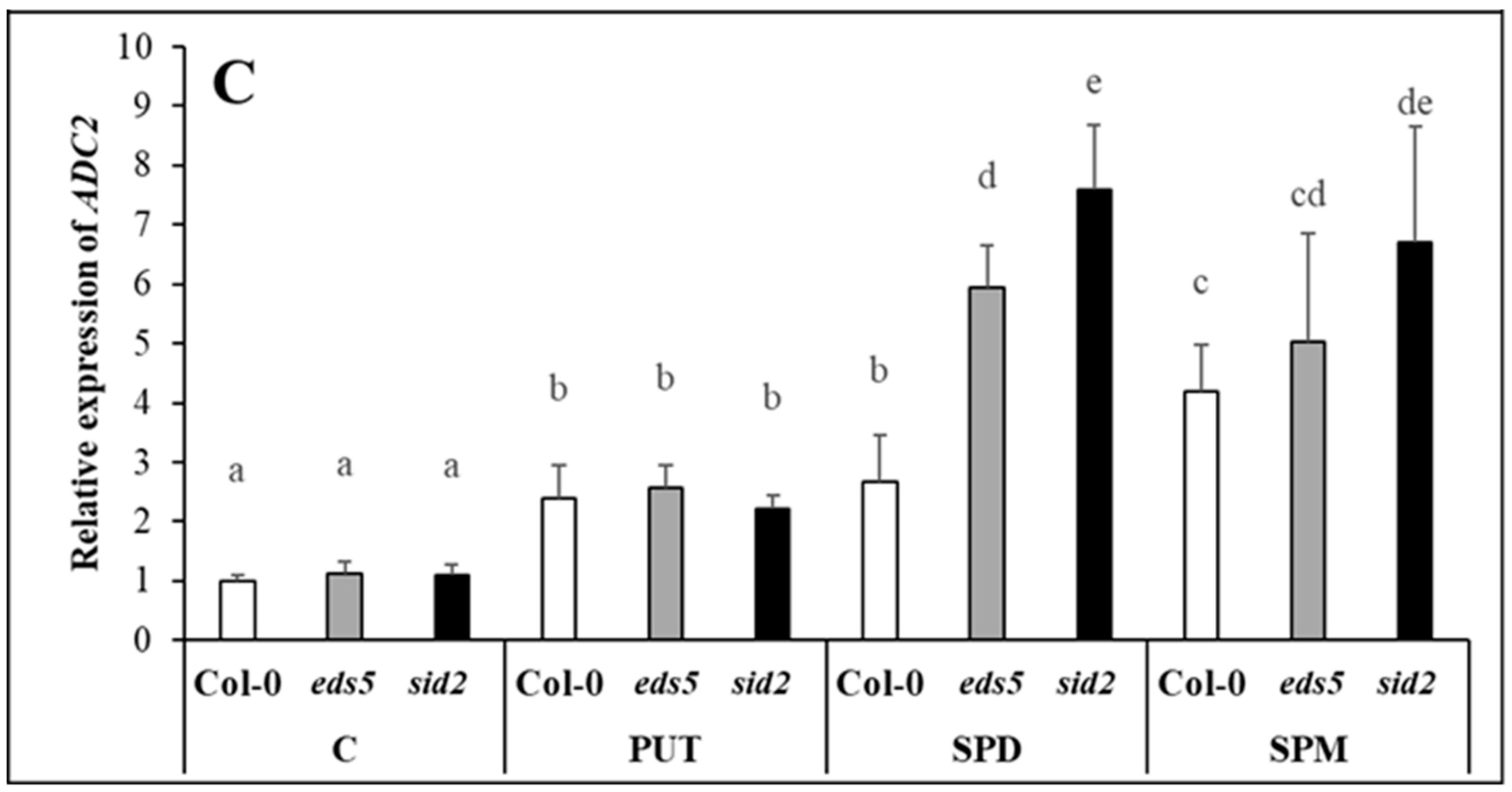
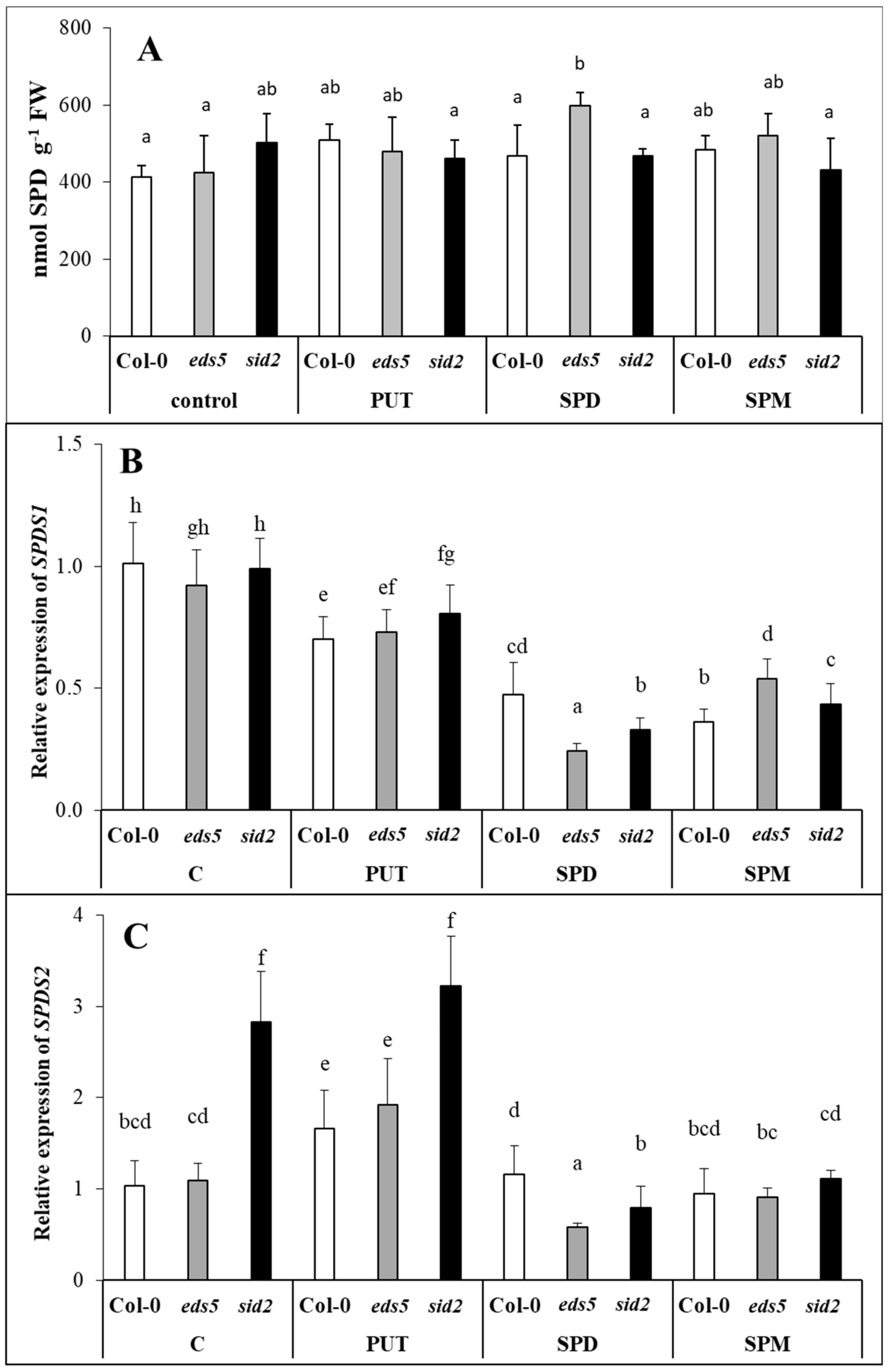
Parallel with these, although the level of higher PAs did not change in the leaves of the plants, the
SPDS1 expression level decreased after SPD or SPM treatments, suggesting that higher PAs, uptaken by the roots is probably not transported to the leaves, inducing the down-regulation of the SPD synthesis in the leaves. A decrease in
SPDS1 expression together with
CuAO1 expression in the SPD- and SPM-treated plants may be responsible for the maintenance of optimal SPD content. It has been reported that significantly induced expression of
CuAO1 was observed after ABA treatment in
Arabidopsis [
43]. In the present experiment, the highest
CuAO1 transcript level was detected parallel with the highest ABA content in the SPD-treated plants. Interestingly, under control conditions and after PUT treatment, the
SPDS2 transcript level was higher in the
sid2 mutant than in the wild type, but this difference disappeared in SPD- or SPM-treated plants, as the application of higher amounts of PAs inhibited its expression in this mutant. The up-regulation of
SPMS was observed after all the PA treatments, which is understandable in PUT- and SPD-treated plants, where in order to decrease the uptaken excess of PUT or SPD, further synthesis was needed to SPM. Despite of these, the SPM content did not show significant increment in either of the SPM-treated plants after 1 day of treatment.
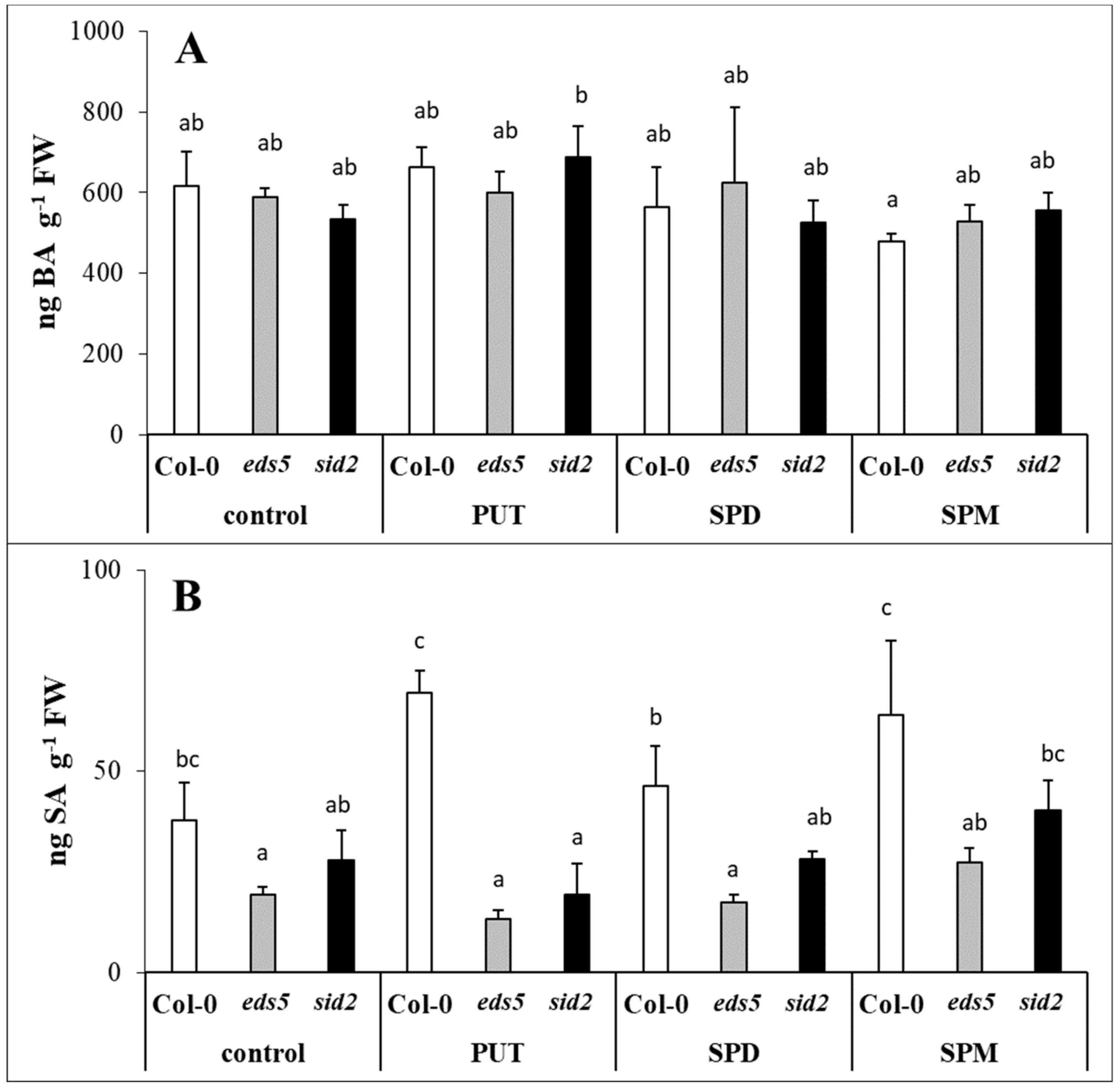
In the present experiment, the SA level was lower in the
eds5 and
sid2 mutants compared to the wild type. As
EDS5 encodes a membrane protein, located at the chloroplast envelope and responsible for SA transport [
44,
45], in the
eds5 mutant, the accumulated SA is trapped in the chloroplast, which in turn can inhibit
ICS expression. While in
sid2 mutants, the SA accumulation can be only a fraction of that of the wild type, because of the lack of the ICS enzyme/pathway. Different concentrations of SA have been reported to have different effects on PA metabolism [
21,
46]. In addition, PA treatments, both as seed soaking or applied hydroponically efficiently enhanced SA content in wheat or maize [
15,
16,
47]. Under the present conditions, PUT and SPM treatments after 1 day could only cause a slight, but statistically not significant increase in SA content of Col-0 genotype. Although,
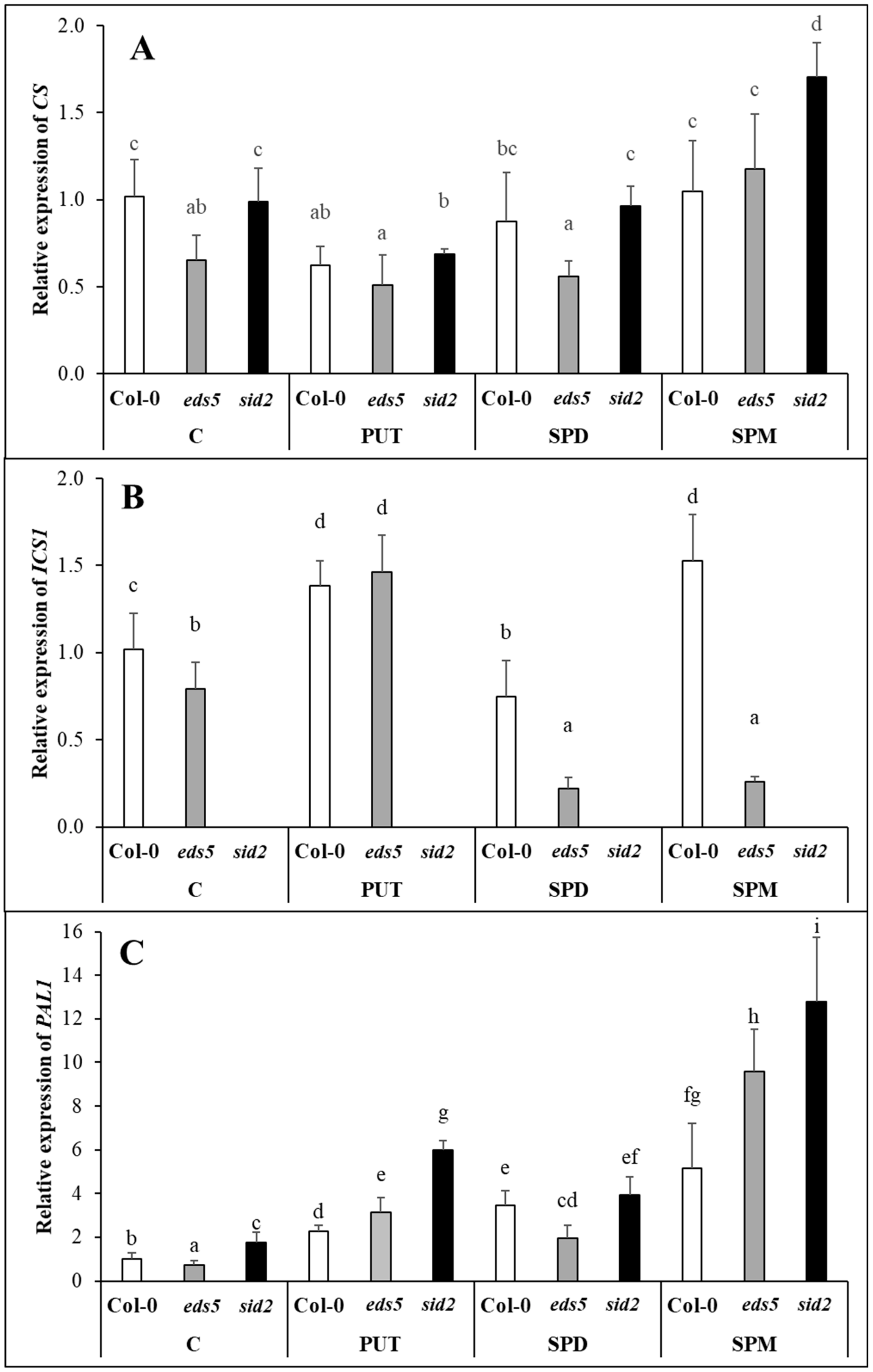
CS expression did not change after either of the PA treatments, the
ICS transcript level in Col-0 was in correlation with the changes in SA content, as it increased after PUT or SPM treatments. Interestingly, PUT also increased the
ICS expression in the
eds5 mutant, but SPD and SPM treatments decreased it. The latter was probably due to the inhibitory feedback effect of the SA accumulation in the chloroplasts, which was not manifested in statistically significant increase in the total SA content. The
PAL1 expression, maybe in order to compensate the mutation of the
ICS1, showed slightly higher basal level in
sid2 plants. Although, PA treatments induced
PAL1 expression in all the genotypes and its up-regulation was the highest in SPM-treated plants, in case of all the treatments it was the highest in the
sid2 mutant. The activity of PAL, a crucial enzyme in the synthesis of flavonoids, anthocyanins and simple phenolic acids, increased after PUT treatment in the leaves of maize [
15]. In addition, in
Atpao4 plants with increased SPM content, genes involved in flavonoid and/or lignin biosynthesis, such as
PAL1 were induced [
48], suggesting that the accumulation of PAs increased the synthesis of phenolic compounds. According to the present results, PA treatments differently induced the SA synthesis pathways. In the Col-0 both pathways activated, especially in PUT- and SPM-treated ones. In the
eds5 mutant, after PUT treatment both pathways induced, and after SPD and SPM treatments the expression level of
PAL further increased, but that of
ICS1 decreased. In the case of
sid2 mutant, the drastic increment in
PAL1 transcript level will be responsible for SA synthesis.
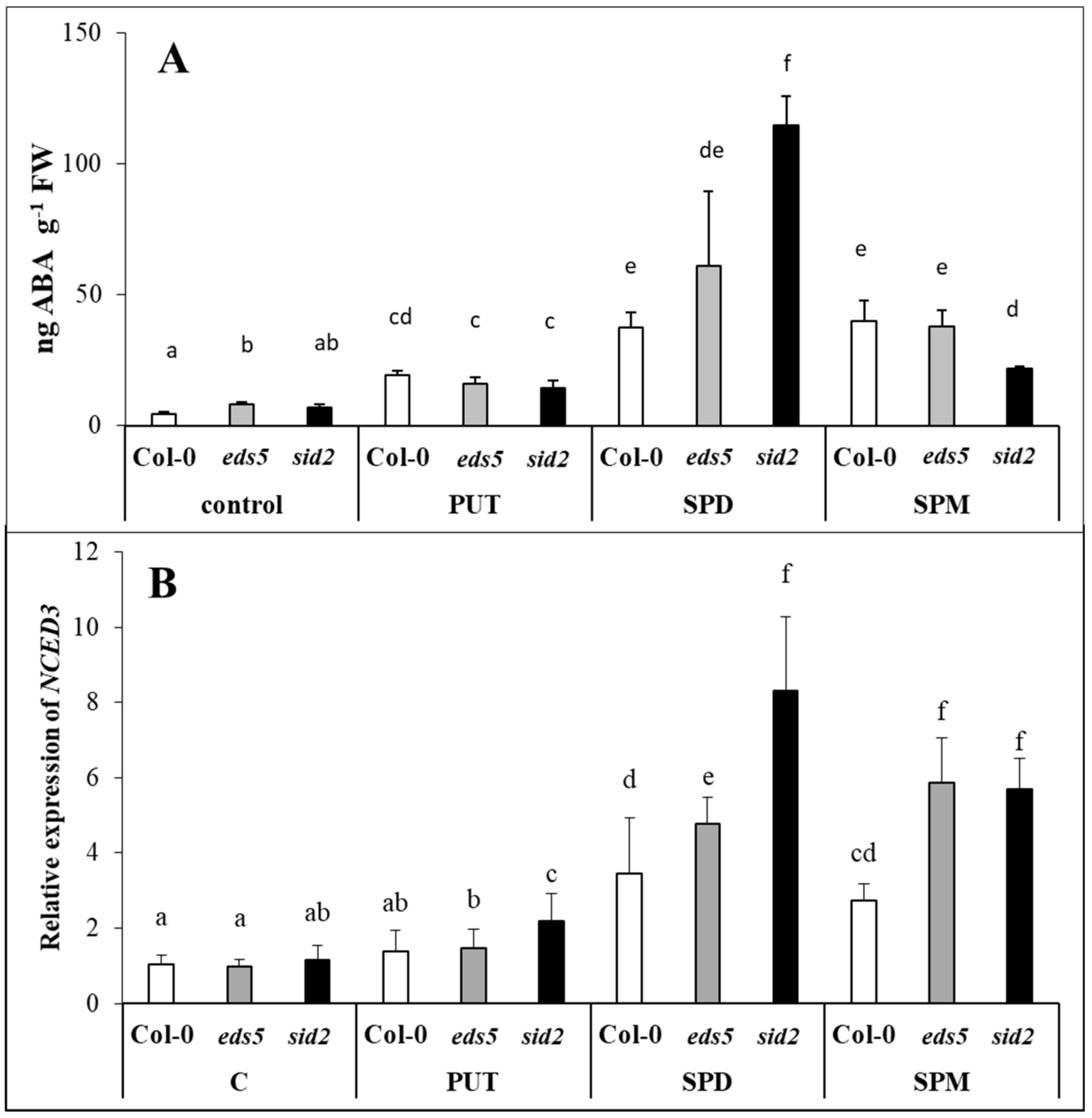
PUT and ABA are integrated in a positive feedback loop [
18]. Modulation of PA metabolism at transcriptional level by ABA has been proved in
Arabidopsis in case of PA biosynthesis genes, such as
ADC2,
SPDS1 and
SPMS [
49]. The transcriptional regulation
NCED in PA-overproducer plants has been demonstrated [
17,
50,
51]. Conversely, the suppression of
ADC resulted in the reduced expression of
NCED and the down-regulation of ABA-regulated genes [
30]. In the present experiment, PAs induced ABA accumulation and the up-regulation of
NCED, which was the most pronounced in SPD-treated plants, where it was the highest in
sid2 mutant. This can be resulted from the well-known antagonistic relationship between SA and ABA [
27,
52,
53]. ABA has been also reported to influence the catabolism of PAs, as its exogenous application induced the expression of
CuAO1 in
Arabidopsis [
42,
43], and
PAO in
Medicago sativa [
54] and wheat [
29]. Interestingly, under the present conditions, the highest
CuAO1 expression was detected in the SPD-treated plants, where the highest ABA accumulation and
NCED expression was found. Nevertheless, all these parameters were the highest in the
sid2 mutant plants. Similarly, higher ABA content has been also detected in
sid2 Arabidopsis mutant compared to the wild type during the transition from pre-reproductive to reproductive stages [
55], confirming that SA content may influence, at least to some extent, the endogenous concentrations of ABA.
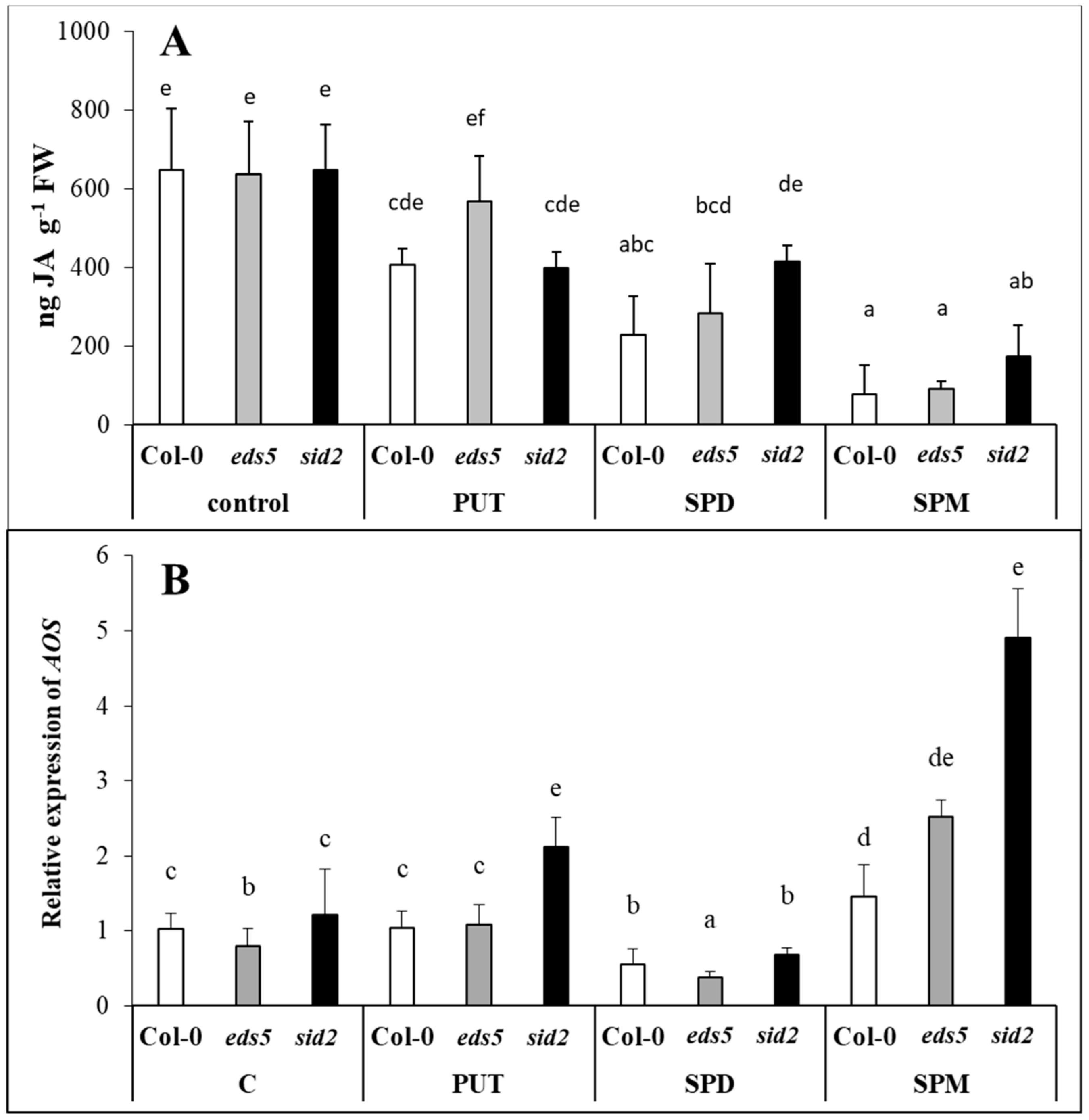
Similar JA content was detected in
sid2 and Col-0 genotypes despite of the different SA level under control conditions and after infection with
Pythium irregular, indicating that SA did not inhibit the JA accumulation [
55,
56]. In the present experiment, different SA contents were also detected parallel with almost the same JA level in the three genotypes. All the PA treatments decreased the JA content, with the highest decrement in SPM-treated plants, but regardless of the genotypes. Exogenous methyl jasmonate has also been reported to increase the production of conjugated PAs in barley [
33]. In addition, genes encoding enzymes for synthesis of PAs, such as ADC1 and SPDS1 have been proved to be affected by JA signalling in
Arabidopsis [
57], and exogenous JA induced PAO in rice [
34], and regulated the expression of chickpea
CuAO [
58]. However, there are only a few studies about the effects of PAs on the synthesis of JA. PA treatments, especially SPM elicited the biosynthesis of JA in lima bean [
35], while constitute overexpression of
SPMS in
Arabidopsis increased the levels of expression of genes involved in JA synthesis and signalling [
17,
59]. Under the present conditions, it was found that all the PAs induced the expression of
AOS, with the highest level in SPM-treated plants, especially in the SPM-treated
sid2 mutant. The
Arabidopsis AOS promoter was found to be activated by a variety of signals, including JA, wounding, and even exogenous SA [
60], however, SA-induced repression of the JA-signalling pathway is independent of JA biosynthesis, as occurs downstream of JA perception [
61]. Despite this, in
eds4 mutant
Arabidopsis (also with reduced SA content) relieved inhibition of JA-dependent signalling responses were detected, confirming that the SA signalling and JA signalling can be mutually inhibitory [
62].
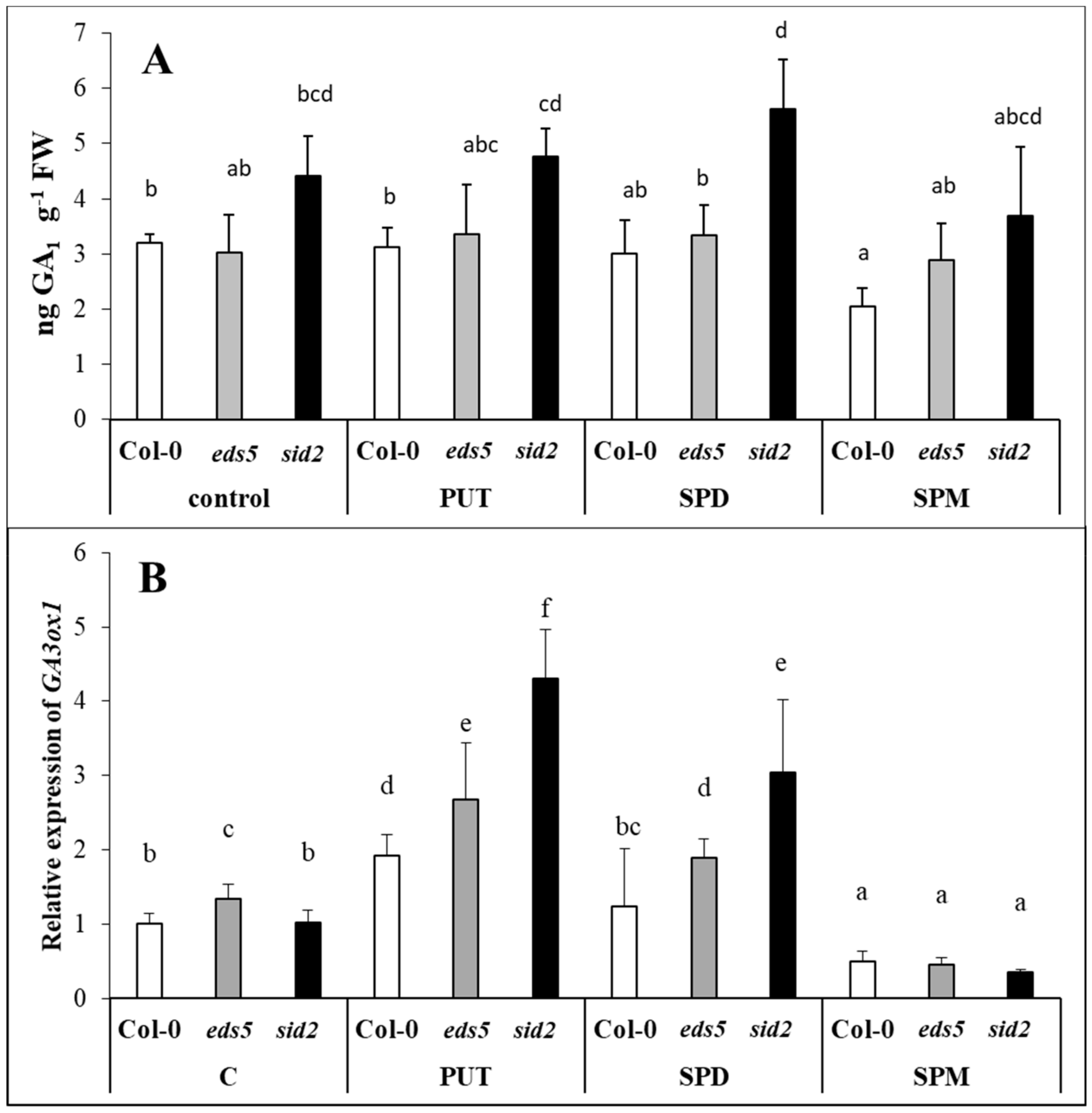
ADC2-overexpressing transgenic plants exhibited a reduction in both the contents of GA
1, 4, 9 contents, and in the expression levels of the
GA20ox1,
GA3ox1 and
GA3ox3 transcripts [
63], suggesting that PUT accumulation represses GA synthesis. SPD treatment decreased GA
3 content, and
GA3ox expression in apple terminal buds during floral induction [
4]. However, on the other hand, an increased content of GAs was observed in PA-treated plants under drought conditions in cree** bentgrass [
64]. SPD increased GA
3 content in maize [
65], and in sweet corn seed embryos [
66]. Nevertheless, accumulation or deficiency of PUT, SPD or SPM did not influence the expression of GA signalling gene in transgenic
Arabidopsis leaves or tomato fruits [
67]. These data indicate that the effect of PAs on GA synthesis is highly dependent on plant species and developmental stage. Under the present conditions, PAs treatments did not influence the GA
1 content, except for a slight decrease in SPM-treated wild type. Although, the
GA3ox1 expression increased after PUT treatment, in each of the genotypes, especially in
sid2 mutant, and after SPD treatment in the mutants, with still a higher extent in case of
sid2, a dramatic decrease was observed after SPM treatment in all the genotypes. According to these, no negative correlation was found between the accumulation of PUT and the level of GA
1, while the inhibitory effect of SPM treatment on
GA3ox1 was pronounced in all genotypes. The existence of crosstalk between GAs and SA signalling in
Arabidopsis was also suggested, as GA treatment increased the endogenous levels of SA and the expression of the
ICS1 gene in Col-0, and the transcript levels of the
GA3ox1 gene were greatly elevated by SA treatments [
68,
69]. However, under the present conditions, in each cases GA
1 accumulation was higher in
sid2 mutant than in the wild type, indicating that SA also influences to some extent GA levels in plants. Nevertheless, here, the GA-ABA antagonism should be also taken into consideration, which has been reported during seed development, plant growth and stress responses. Stress-induced increases of ABA level were parallel with decreases in the GA level and the suppression of the GA synthetic enzyme genes in
Arabidopsis [
70,
71].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










