1. Introduction
Glioblastoma (GBM) is the most common primary malignant brain tumor. GBM is associated with poor prognosis and a life expectancy of approximately 15 months despite optimal therapy, which includes surgery, chemotherapy, and radiotherapy [
1]. The lack of therapeutic success has been attributed to a variety of factors, including rapid infiltration of brain tumor cells, inter- and intratumor heterogeneity, limited diffusion of therapeutic drugs across the blood–brain barrier and brain parenchyma/tumor, and the presence of GBM stem cells (GSC) in the tumor, which are resistant to radiotherapy and chemotherapy and are capable of tumor formation and indefinite self-renewal [
2].
Graphene is made up of a layer of carbon atoms arranged in a hexagonal pattern and consists purely of sp2 hybridized bonds. It has gained enormous interest in various fields owing to its unique electrochemical properties, which include high thermal conductivity, high current, density, chemical volume, optical transmittance, and very high hydrophobicity [
3,
4]. It is the simplest form of carbon and the thinnest material ever produced [
5]. The graphene family includes sheets and flakes of graphene as well as graphene oxide (GO) and reduced graphene oxide (rGO) [
6,
7]. GO is highly hydrophilic because of the presence of a large number of oxygen groups on the surface (hydroxyl, carboxyl, epoxy). rGO has more oxygen functional groups than graphene (GN), but less than GO. Therefore, rGO is less hydrophilic than GO and, therefore, has higher electrical conductivity [
8]. In addition to the numerous applications of graphene materials in electronics, their derivatives are also assessed for their use in medicine, for example, in anticancer therapy [
9,
10]. Our previous studies have shown that graphene and its derivatives can be cytotoxic to glioblastoma cells in vitro and in vivo. GN, rGO, and GO induce apoptosis and lead to the reduction of viability and proliferation in U87 and U118 glioblastoma cell lines [
11]. Jaworski et al. also showed that graphene flakes were too large (0.45–1.5 µm) to enter the glioblastoma cells [
11,
12]. However, both GN and rGO can activate the mitochondrial-dependent apoptotic pathway by reducing the mitochondrial membrane potential in U87 glioma cells. On the contrary, GO can regulate the expression of mitochondrial oxidative phosphorylation (OXPHOS) genes in GBM, thus leading to a decrease in the invasion potential of cancer cells [
13]. The tested materials (GN/exfoliation (ExF), rGO/thermal (Term), rGO/ammonium thiosulphate (ATS), and rGO/ thiourea dioxide (TUD)) showed no effect on the cell cycle [
14]. Despite the fact that even minimal oxygen content on the surface of the flakes may reduce the proapoptotic abilities of graphene and its derivatives, oxygen-containing derivatives have a better affinity for GBM cells than pure graphene, which allows for better targeting of the intended effect [
15].
An important finding seems to be the reduction of oxygen content in rGO in relation to the starting material, GO, which results in an increase of delocalized electrons on the surface of the graphene flakes. This can lead to a disruption of the signaling pathways in the plasma membrane or to direct interaction with cell structures, which are sensitive to electrochemical potential. Therefore, graphene and its derivatives are characterized by strong bioelectric properties; because of the presence of delocalized electrons and oxygen-containing functional groups on the surface of the flakes, this material can interact with structures that receive electrical signals, for example, receptors located on the cell surface, as well as proteins building voltage-dependent ion channels, ultimately causing changes in the potential of the cell membrane. Cell surface charge is a key biophysical parameter that depends on the composition of the cytoplasmic membrane and the physiological state of cells. In addition to the presence of ion channels and transporters, negative values of cell membrane potential at physiological pH values are also caused by the presence of nonionic groups in phospholipids (phosphatidylcholine; −62 mV), proteins, and their polysaccharide conjugates [
16]. Direct comparisons of the in vitro and in vivo Vm levels of normal and cancer cells showed that the cancer cells were more depolarized (negative Vm) than their normal counterparts [
17]. Therefore, the studied materials of graphene origin (GN and rGO) may prefer adhesion to neoplastic cells with depolarized cell membranes. Moreover, Fiorillo et al. proved that GO, characterized by the presence of many functional groups containing oxygen, preferred adhesion to U87 tumor cells rather than stem cells and normal fibroblasts [
18]. Therefore, it has been hypothesized that graphene-derived flakes exert modulating and transducing effects on proteins contained in the membrane of U87 glioma cancer cells. These effects result from the unique structure and type of functional groups on the surface of graphene. GN flakes are characterized by several functional groups on the surface, and they possess a larger electron cloud than rGO.
Ion channels play an important role in the regulation of electrical excitability in normal and cancer cells [
19]. Ion channels and transporters are also associated with GBM tumor growth and malignancy [
20]. Genomic analysis revealed that the genes involved in Na, K, Ca transmission or transport belong to the most frequently mutated functional groups affecting GBM in 90% of the tested samples [
21,
22]. Therefore, the article discusses the subject of studying the expression of individual subunits that build ion channels (Na
+, K
+, Ca
2+, Cl
−). The functioning of ion channels and pumps influences the migration and proliferation of GBM cells. For example, deregulated K
+ and Cl
− channels regulate the osmotic drive, allowing for cell shape and volume changes that promote glioblastoma cell migration [
23], and Ca-activated K
+ (BK) channels, which control glioblastoma cell growth [
24]. In this study, we hypothesize that flakes of graphene (GN) and reduced graphene oxides (rGOs) may affect the expression of voltage-dependent ion channel genes Cl
− (
clcn3 and
clcn6), Ca
2+ (
cacna1b and
cacna1d), Na
+ (
nalcn), K
+ (
kcne4,
kcnj10, and
kcnb1), resulting in an alteration of the potential of the glioblastoma cell membrane. It is also suspected that GN and rGO flakes may reduce the expression of extracellular receptors and further reduce the invasiveness of glioblastoma in vitro.
The aim of this study is to determine changes in the cell membrane caused by direct or indirect contact with bioelectric flakes of rGO with different degrees of reduction compared to GN in U87 glioblastoma multiforme cells in vitro.
3. Discussion
Previous studies have shown that reduced graphene oxides (rGOs), compared to graphene GN, does not exert identical effects on glioblastoma multiforme cancer cells. The effectiveness of the tested materials was examined in terms of viability, metabolic activity, cell cycle dynamics, and the level of apoptosis. The results indicate that GN and rGOs activate the mitochondria-dependent apoptotic pathway by reducing the potential of the mitochondrial membrane. This study has proved that rGOs have a stronger cytotoxic effect than GN. We used the MTT assay to assess whether graphene and its derivatives also affect the viability and metabolic activity of fibroblast Hs5 and glioblastoma U87 cells. The highest decrease in metabolic activity in glioblastoma U87 cells, at 8.69% ± 12.88%, was found in the group treated with rGO/TUD at a concentration of 100 μg/mL. Interestingly, in the group treated with rGO/ATS, the lowest viability, at 37.7% ± 12.55%, occurred at a concentration of 5 μg/mL rGO/ATS. In other groups treated with GN/ExF and rGO/Term, ~50% mortality was observed at concentrations ranging from 25 to 100 μg/mL. Reducing the oxygen content and increasing defects in the connections between carbons in rGOs compared to GO resulted in an increase in the number of delocalized electrons on the surface of the graphene flakes and oxygen groups, including hydroxyl, carboxyl, and epoxy [
14]. It was hypothesized that the cytotoxicity of rGOs may mainly result from direct contact with the glioblastoma cell membrane and may lead to the disruption of signaling pathways in the plasma membrane or direct interaction with cell structures, which are sensitive to electrochemical potential (cell membrane, e.g., ion channels and extracellular receptors). Studying the interactions between graphene materials and cell membranes may reveal the underlying mechanisms of the cytotoxicity of these materials.
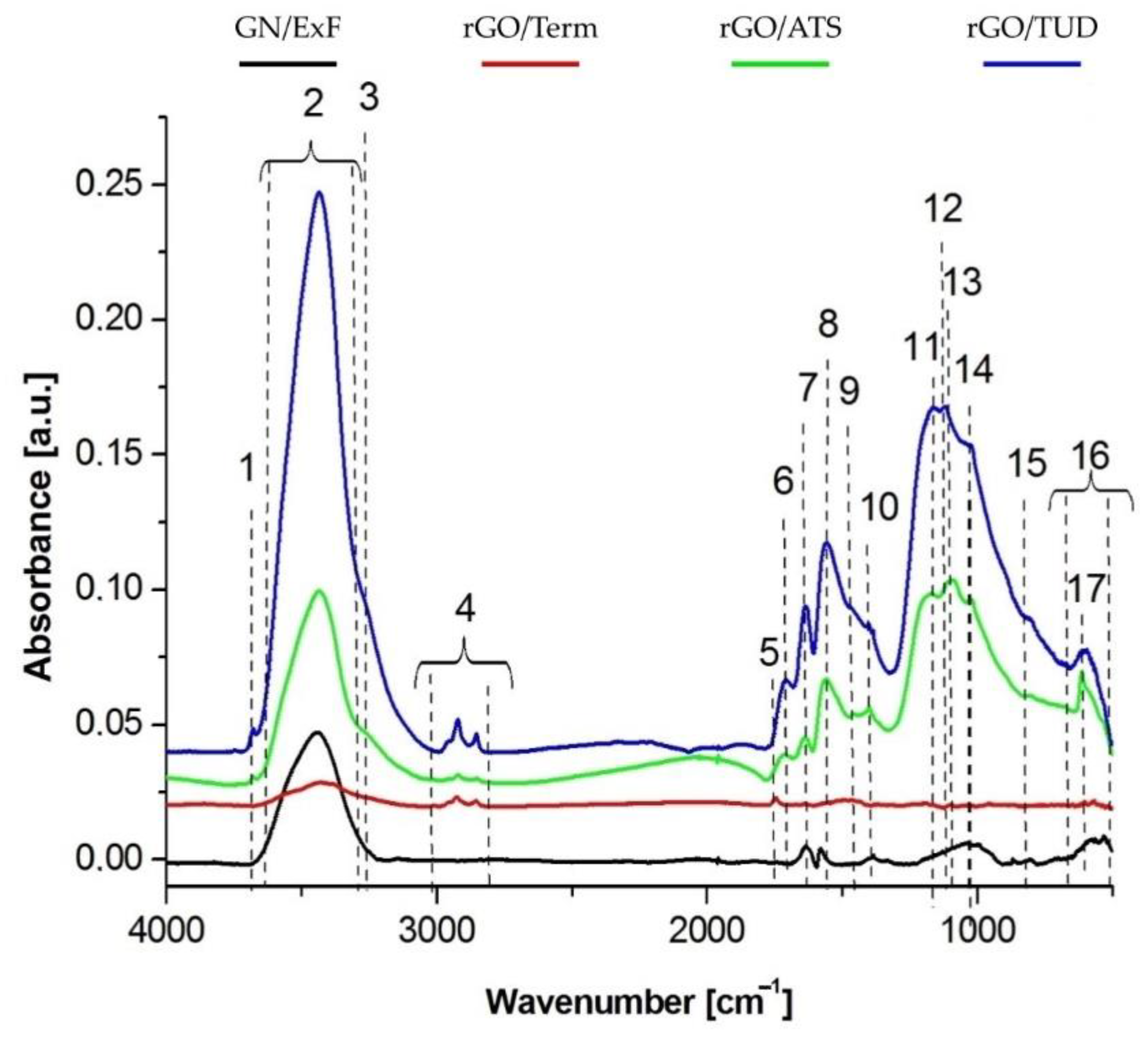
Using FTIR analysis, a detailed examination was performed to compare the presence of characteristic functional groups in both reduced graphene oxides (rGO/Term, rGO/ATS, rGO/TUD) and graphene (GN/ExF). This analysis confirmed the results obtained in a previous study [
14]. The FTIR analysis presented in this paper shows the presence of hydroxyl groups (-OH) on the surface of the flakes, derived from the native aqueous solution in which the flake suspensions were prepared. Further, the presence of C=O connections may be associated with a greater affinity of the material for the attachment of hydroxyl (OH-) groups. Thus, the more C=O bonds that are hydrophilic in nature, the more OH- hydroxyl groups. The presence of carbonyl groups (C=O) and carboxyl groups (COO-) was also observed. The highest expression was observed in the rGO/TUD treatment. In a study by Loryuenyong et al., the presence of hydroxyl (-OH), carbonyl (C=O), and epoxy (C-O) groups was confirmed in GO and rGO. Moreover, the high intensity of the main peaks in GO confirms the presence of a large number of oxygen functional groups after the oxidation process [
25]. Emiru and Ayele obtained a similar band arrangement of the FTIR spectrum of GO and rGO, as presented in this article. Among other things, they observed such functional groups as OH-, COH-, COOH-, and CO- [
26]. It showed that with the use of chemical methods, the content of functional groups is reduced. The most effective method for GO reduction is the thermal process because the obtained rGO/Term had the highest rate of reduction [
14].
It is known that graphene materials can modulate electron transfer in redox reactions. The speed of the redox reactions on the graphene surface depends on the migration of electrons across the reactive surface and the subsequent transfer along the surface. Pan et al., during an analysis of the resistance in the studied graphene materials, proved that the higher the degree of graphene folding, the more difficult it was to transfer electrons [
27]. The same relationship between the degree of folding and resistance was observed in a previous study [
14]. Thus, the more corrugated the surface of the carbon material (rGO/Term < rGO/ATS < rGO/TUD), the lower the resistance of the graphene flakes.
The obtained selectivity of graphene flakes towards cancer cells could be determined by the differences in cell membrane potential (Vm) between cancer and noncancerous cells. Cell surface charge is a key biophysical parameter that depends on the composition of the cytoplasmic membrane and the physiological state of cells. Cone’s theory [
28] proposes a general correlation between proliferation and Vm, as he showed the significant depolarization of Vm during the malignant transformation of normal cells [
29,
30]. Direct comparisons of the in vitro and in vivo Vm levels of normal and cancer cells showed that the cancer cells were more depolarized than their normal counterparts [
17]. A cell depolarizes when Vm is relatively less negative (tumor cells, proliferating: 0 to −50 mV), while a hyperpolarized cell has more negative Vm (normal cells, nonproliferating; −50 to −90 mV) [
31]. Glioblastoma cells express the potential of −14 mV of depolarized cell membrane, which determines the lower repulsive forces. Therefore, the flakes with reduced graphene (rGO/ATS and rGO/TUD in particular) possess a lower negative surface charge and can probably adhere to glioblastoma U87 cell membranes more easily.
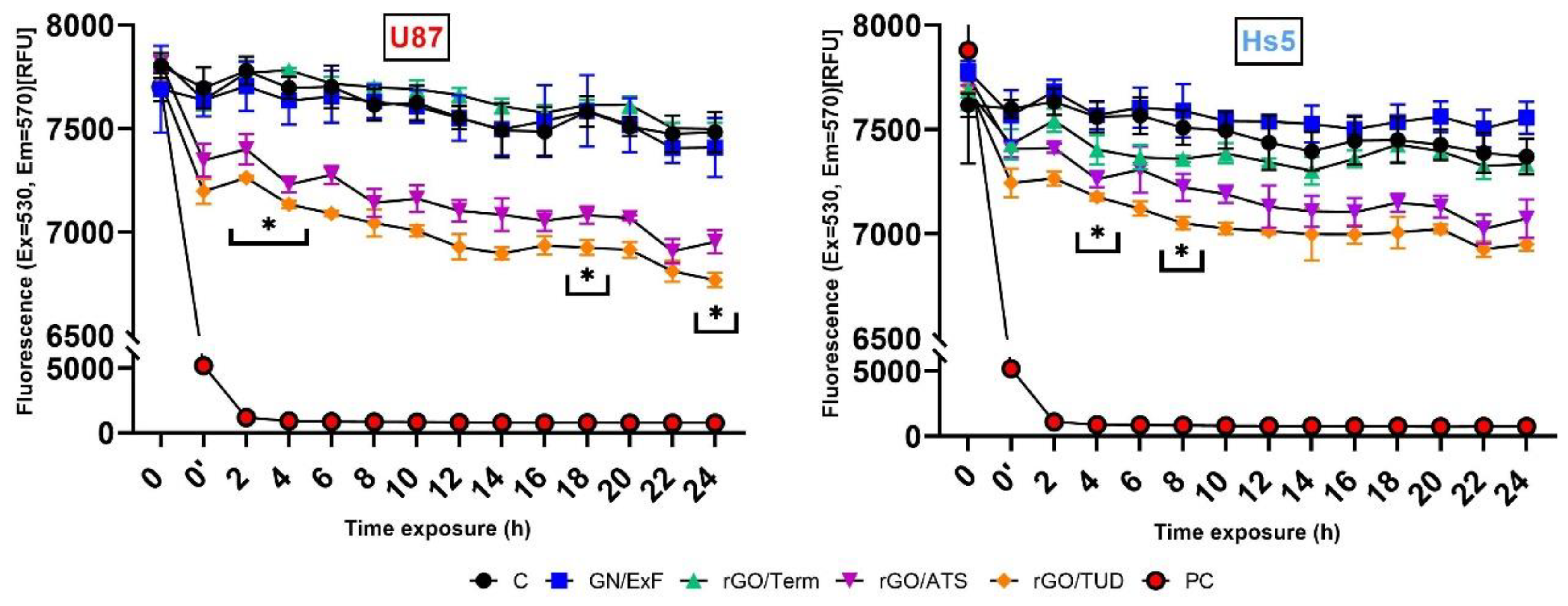
When analyzing the results of the cell membrane potential (Vm) of U87 and Hs5, a decrease in cell membrane potential was observed over time after treatment with rGO/ATS and rGO/TUD flakes. However, the decrease in potential in fibroblast Hs5 cells was smaller than that in U87 cells: for rGO/ATS material, it was 258.4 RFU and 421 RFU, respectively, and for rGO/TUD material, 384.3 RFU and 561.7 RFU, respectively. Bondar et al. observed that the membrane potential in HeLa cells, in which apoptosis was thermally induced, shifted negatively by about 4.2 mV compared to control cells. This was probably the result of the redistribution of phosphatidylserine, containing a negatively charged carboxyl group, from the inner to the outer lipid layer of the cell membrane [
16]. Therefore, the reduction of the U87 glioma cell membrane potential after treatment with rGO/ATS and rGO/TUD flakes may be partially secondary to the flakes due to the induction of apoptosis in cells.
Graphene flakes, apart from direct contact with the lipid double membrane, are in contact with channels, including ion channels. Ion channels are membrane proteins that open or close a plasmatic membrane, depending on a voltage gradient or ligand binding. They are essential for cell proliferation and play a key role in malignant glioma by influencing the shape and volume of glioblastoma cells, which, in turn, may influence the invasiveness and migration of tumors [
32].
The effect of the graphene flakes on the expression of voltage-gated ion channels, participating in both electrical and chemical signaling pathways [
33], was assessed. Wang et al. showed that between 18 genes of ion channels (voltage-gated and ligand-gated), the expression of 16 genes (
cacna1d,
clcn6,
glrb,
gria2,
grid1,
kcnab1,
kcnb1,
kcnd2,
kcnj10,
kcnma1,
kcnqn3,
nalcn,
p2rx7,
scn1a, and
vdac2) was reduced compared to normal tissue [
34]. Based on these studies, genes encoding voltage-dependent ion channel subunits (
cacna1d,
clcn6,
kcnab1,
kcnb1,
kcnj10, and
nalcn) were selected to determine the effect of graphene derivatives on channel gene expression. The following genes were also added for the analysis:
clcn3 [
35],
cacna1b [
36], and
kcne4 [
37].
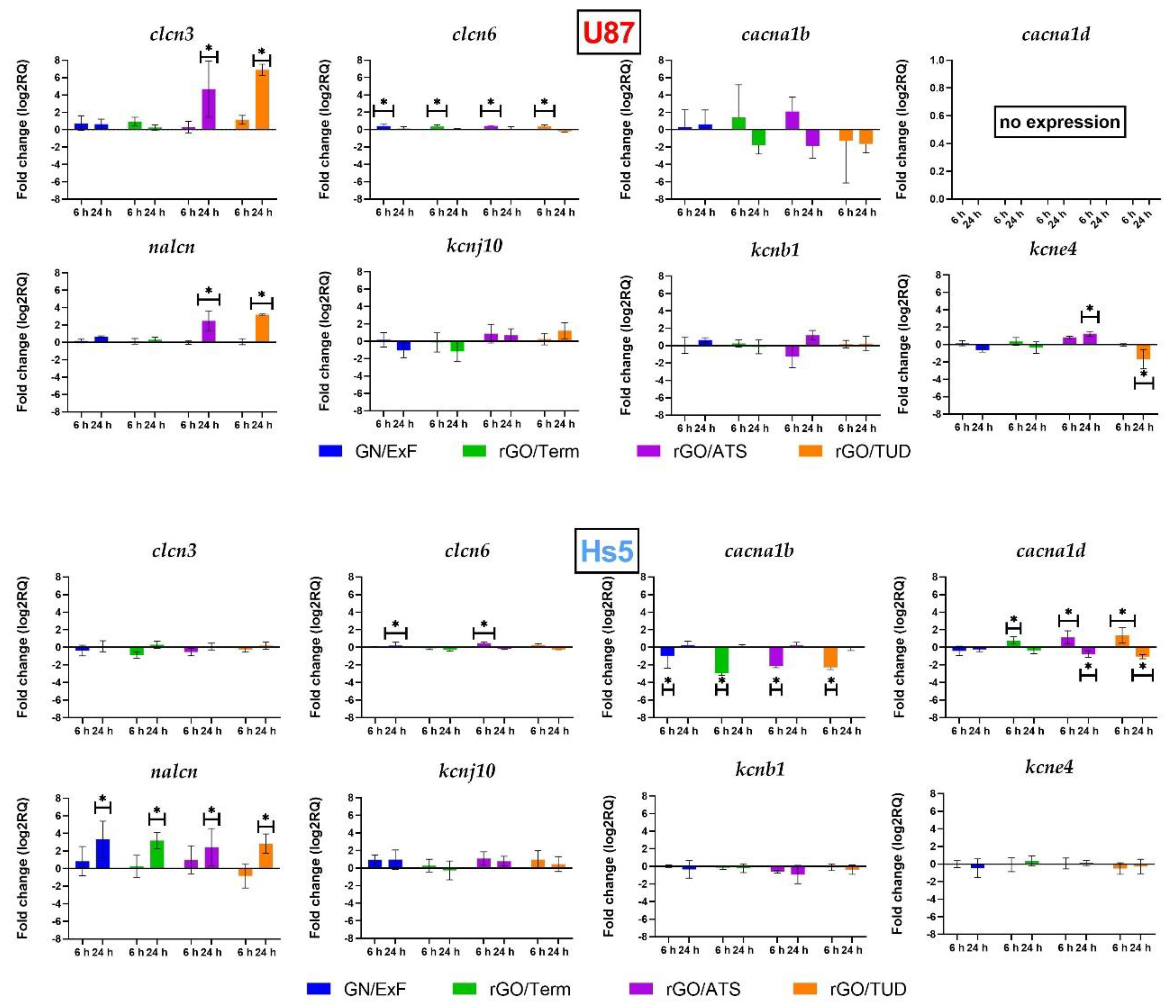
The presented study shows that the expression of
nalcn in U87 glioma tumor cells was significantly higher (log2RQ = 6.15) than in Hs5 fibroblasts. Ouwerkerk et al. reported that the concentration of Na
+ ions in malignant tumors increased in comparison to noncancer tissues [
38]. Moreover, the expression of
nalcn in astrocytes, the glial cells that glioblastoma is derived from, was markedly low [
39]. The expression of
nalcn in U87 glioma cells was significantly higher after treatment with rGO/ATS and rGO/TUD. In Hs5 cells, the same effect was observed in the case of each treatment with graphene derivative material. It was suggested that the increase of intracellular Na
+ occurs in the early phase of apoptosis [
40,
41]. Moreover, several studies also reported an elevation of cytoplasmic Na
+ in the late phase of apoptosis [
42,
43]. Thus, an increased expression of
nalcn, induced by the graphene flakes, can provoke apoptosis in glioblastoma cells by stimulating sodium influx into the cells, which increases the cytoplasmic concentration of Na
+.
clcn6 is predominantly localized in the intracellular vesicles of the endoplasmic reticulum (late endosomes) and the cell membrane. Our study showed a significant increase of
clcn6 during the initial treatment of U87 glioblastoma cells in all tested groups. On the contrary, Hs5 cells revealed a significant increase of
clcn6 expression in the GN/ExF and rGO/ATS treatment groups. The observed overexpression leads to lysosomal acidification [
44]. Neagoe et al. clearly proved that CIC-6 mediates the Cl
−/H
+ exchange, which affects its coupling, conductivity, and ion selectivity features [
45]. Increased expression of the
clcn6 gene in U87 glioma cells leads to cytoplasmic alkalization. In fact, intracellular alkalization and extracellular acidification are commonly observed in malignant tumors. The altered activity of cell transporters, internal enzymes, and pH gradient in the cancer cell membrane plays a pivotal role in tumor progression and metastasis [
46,
47]. Acidification of the cytoplasm leads to the activation of apoptotic pathways in cancer cells [
48]. Based on that, we conclude that graphene flakes (GN/ExF) and rGOs in U87 glioblastoma cells block Cl
−/H
+ transport. Consequently, it leads to the acidification of the cytoplasm and the activation of apoptosis. Simultaneously, in response to that blockage, an increase in
clcn6 gene expression can occur.
Four classes of potassium channels are distinguished: Kv channels (voltage-gated), KCa
2+ channels (calcium-activated), Kir channels (inward-rectifier potassium channel), and K2P channels (two-pore channels) [
49,
50]. Based on these studies, selected markers of Kv and Kir classes were verified. In the channels of the Kv class, we analyzed expression
kcnb1 (Kv2.1) and
kcne4 channels, as well as
kcnj10, which belongs to the Kir potassium channel class. Major changes were observed in the expression of the
kcne4 channel belonging to the Kv potassium channel class. This study confirmed a significant increase in the expression of the
kcne4 channel (log2RQ = 1.192) after 24 h treatment of U87 glioma cells with rGO/ATS. Meanwhile, the reduction in
kcne4 expression (log2RQ = −1.734) was observed after 24 h treatment by rGO/TUD. No changes in KCNE4 expression were noted in Hs5 fibroblasts. KCNE4 expression was characterized by a 2.9-fold increase in glioma compared to the healthy tissues [
51]. The
kcne4 genes encode single proteins of the transmembrane domain with an extracellular
N-terminus and an intracellular
C-terminus. Therefore, the abovementioned proteins cannot form functional ion channels. They can function as the accessory subunits for various ion channels and regulate their biophysical and pharmacological properties in parallel [
52,
53,
54].
The study confirmed no changes in
cacna1b expression in U87 cells after treatment with different graphene materials. In Hs5 cells, the expression was decreased in all experimental groups in the initial treatment. The results confirmed that the expression of the
cacna1b gene in the fibroblasts was significantly higher (log2RQ = 7.52) than in U87 cells. Wang et al. reported, based on brain and breast cancer studies, that
cacna1b is expressed at a low level in tumor cells. Brain cancers, including glioblastoma, oligodendroglia, anaplastic astrocytoma, diffuse astrocytoma, and glioblastoma, show a significant reduction of
cacna1b expression compared to the control groups [
55].
In our studies, we did not observe the expression of the cacna1d gene encoding the subunit of calcium channels in U87 cells. Changes in cacna1d expression occurred only under rGO influence in Hs5 cells. At the initial 6 h treatment, an increase of cacna1d was observed after rGO/Term, rGO/ATS, and rGO/TUD treatment. However, 24 h of treatment decreased its expression in both rGO/ATS and rGO/TUD groups. The primary increase in cacna1d expression can induce a greater influx of Ca2+ ions into the cells. Therefore, after 24 h of treatment, we noticed the reduction in cacna1d expression, which equalized the earlier influx of Ca2+ ions.
Analysis of
clcn3 expression in U87 glioma cells showed an increase in expression after treatment with graphene flakes (GN/ExF) and reduced graphene oxide flakes (rGO/Term, rGO/ATS, rGO/TUD). In particular, significant changes were visible in the rGO/ATS and rGO/TUD groups. Recent studies have shown that
clcn3 is highly expressed in GBM, and it plays a significant role in cell survival, proliferation, and malignancy [
56,
57]. Sontheimer et al. showed that decreased expression of
clcn3 channels inhibits the migration of glioblastoma cells in vitro and in vivo [
35]. An increase in
clcn3 expression can stimulate the invasiveness of U87 glioma cells. No parallel changes were observed in Hs5 fibroblasts after treatment with carbon flakes.
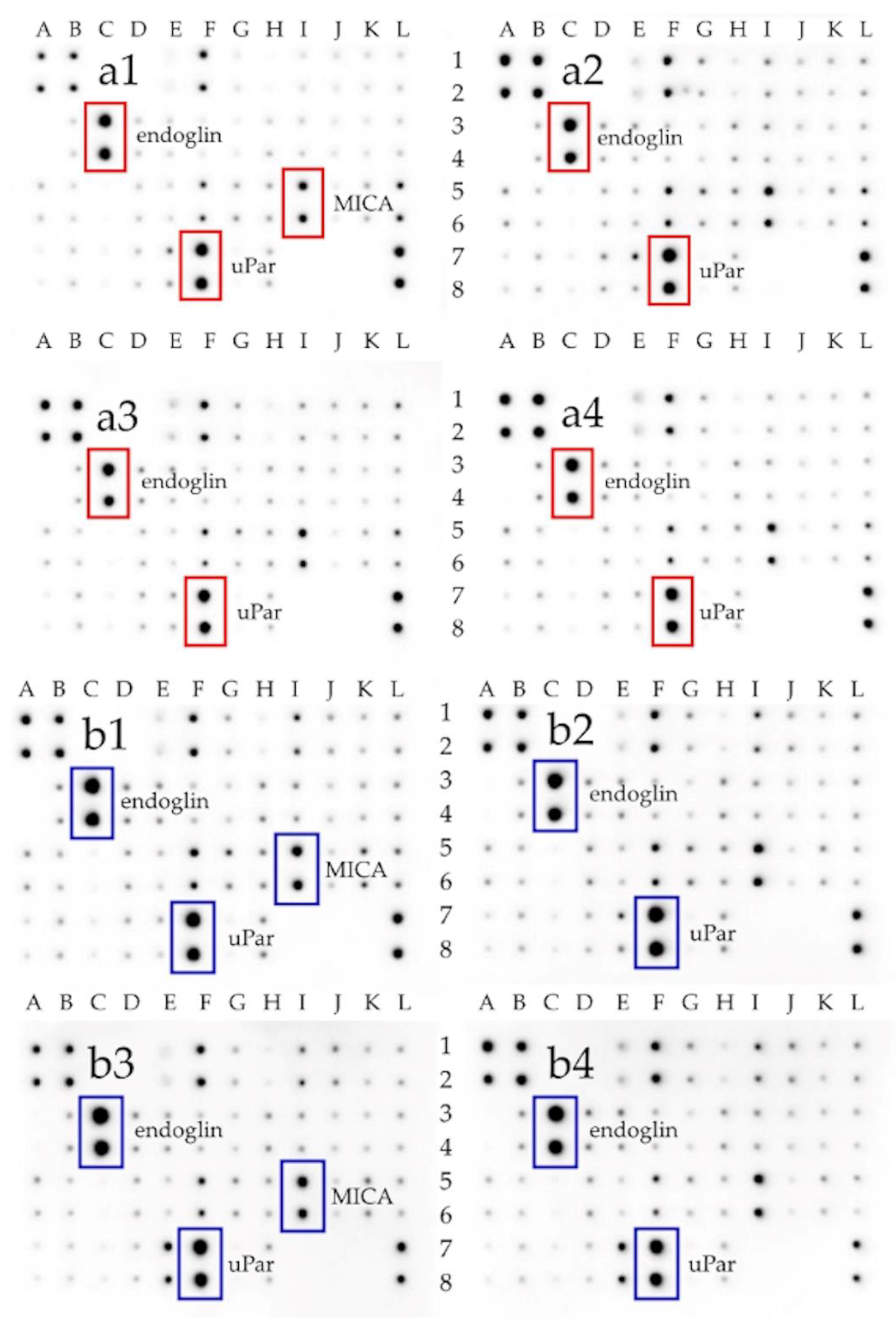
Additionally, using the protein matrix, which allows the determination of the expression of 40 different receptor proteins involved in different signaling pathways (
Figure 5), we assessed the expression of selected extracellular receptors. As a result, it was noticed that the expression of the uPar protein was increased in U87 cells compared to Hs5 cells. Raghu et al. confirmed significantly higher uPar protein expression in U87 cells in comparison to normal HMEC cells [
58]. Analysis of uPar expression in U87 glioma cells after treatment with graphene flakes (GN/ExF) and reduced graphene oxide (rGO/ATS, rGO/TUD) showed a significant reduction of its expression. It was mainly affected by the treatment with reduced graphene oxides. uPar is responsible for the degradation of extracellular matrix (ECM) components attached to the cell surface [
59]. It contains three domains connected to the concave structure, which is the binding site for vitronectin. However, since the binding sites of vitronectin and uPa are distinct, uPar can bind simultaneously to both ligands, regulating proteolysis, adhesion, and cell signaling [
60]. The association between uPa and uPar can cause the cleavage of the adjacent uPar molecules. The cleaved uPar does not support plasminogen, which is mediated by the activation of uPa on the cell surface [
61]. Thus, the flakes of graphene and reduced graphene oxides act similarly to uPa by activating the cleavage of adjacent uPar receptors. They can also block the attachment sites of uPar and vitronectin and, therefore, inhibit uPar signaling [
61,
62].
Endoglin (CD105) is the other factor that had a changed expression after treatment with GN and rGO flakes. It is transmembrane homodimeric protein localized in the endothelial cells of blood vessels. It is a component of the transforming growth factor β (TGFβ) receptor signaling pathway. CD105 plays a pivotal role in angiogenesis and vasculogenesis processes, preventing apoptosis in hypoxic endothelial cells [
63]. It was observed that CD105 is correlated with cancer prognosis (particularly in pediatric cases), but its role in high-grade gliomas remains unclear [
64]. Our study showed a significantly higher endoglin expression in U87 glioma cells than in Hs5 fibroblasts. In other studies, a high expression of CD105 in neoplastic tissue, such as in meningiomas [
65] or childhood brain tumors [
64], was also confirmed. The presented study showed a decrease in CD105 expression in U87 glioma cells after treatment with GN and rGOs in all treatment groups. Muenzner et al. confirmed that the endoglin carboxy-terminal domain is required to inhibit cell detachment [
66]. Therefore, treatment with GN and rGO, resulting in decreased endoglin expression, can stimulate cell adhesion and, consequently, leads to the reduction of the ability of cancer cell migration.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}