
Bioprospecting of Less-Polar Fractions of Ericaria crinita and Ericaria amentacea: Developmental Toxicity and Antioxidant Activity

 , and
, and
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Toxicological Screening of Fractions Obtained from Ericaria Species
Zebrafish Embryotoxicity Test
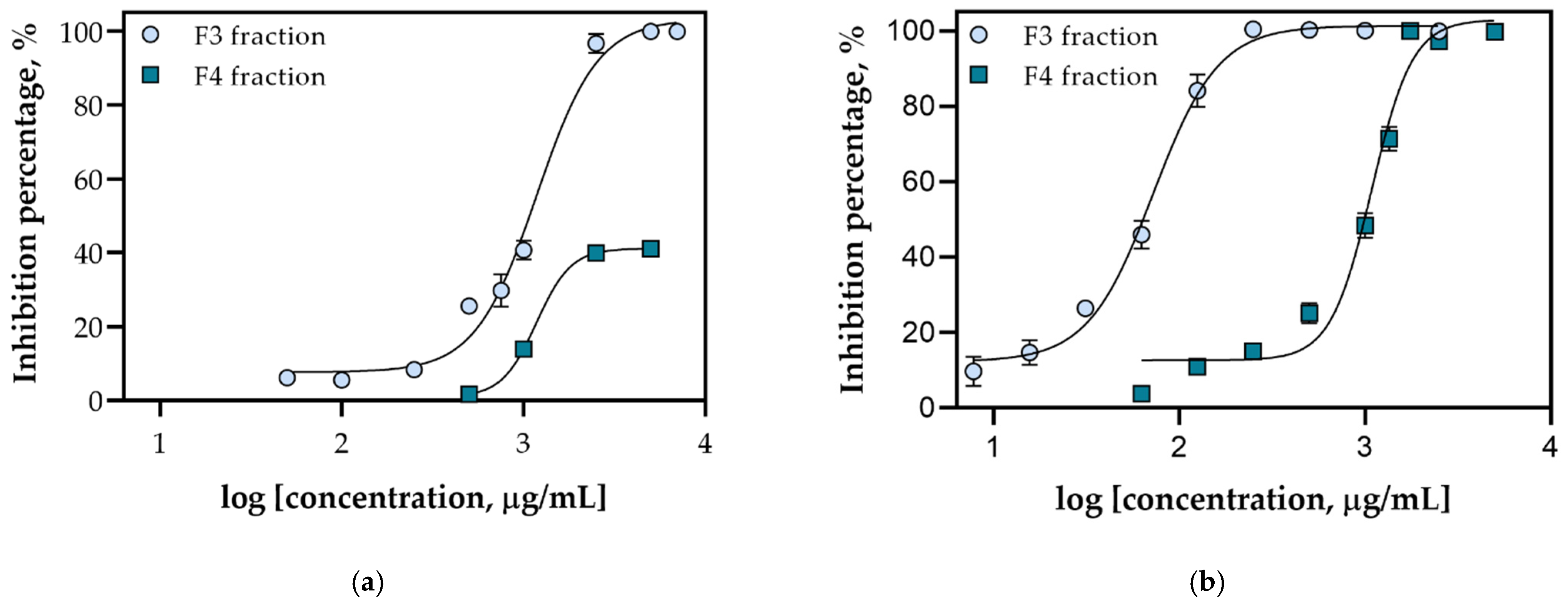
2.2. Screening of the Antioxidant Activity of Fractions Obtained from Ericaria Species
3. Materials and Methods
3.1. Chemicals
3.2. Macroalga Samples
3.3. Fractionation by Solid-Phase Extraction (SPE)
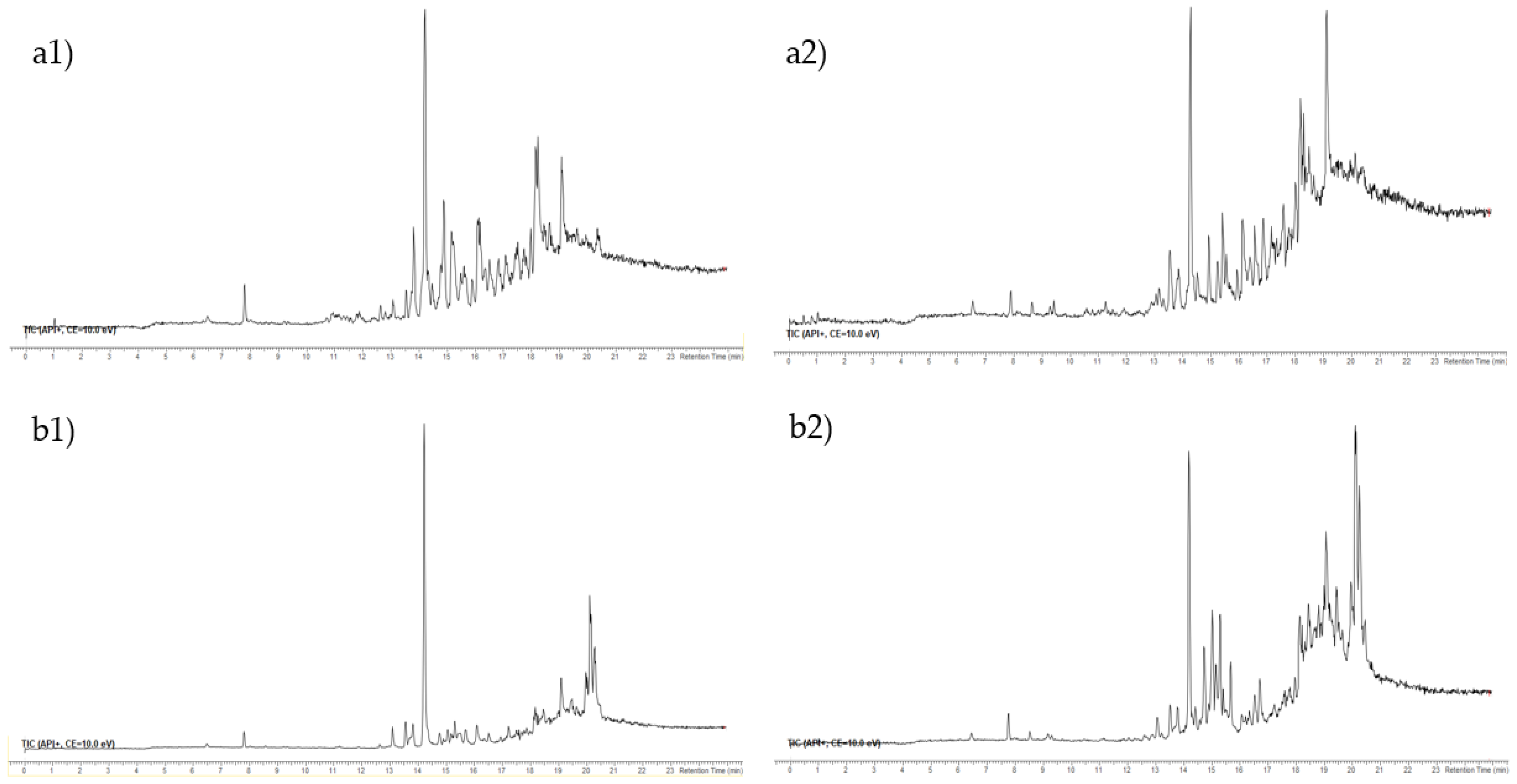
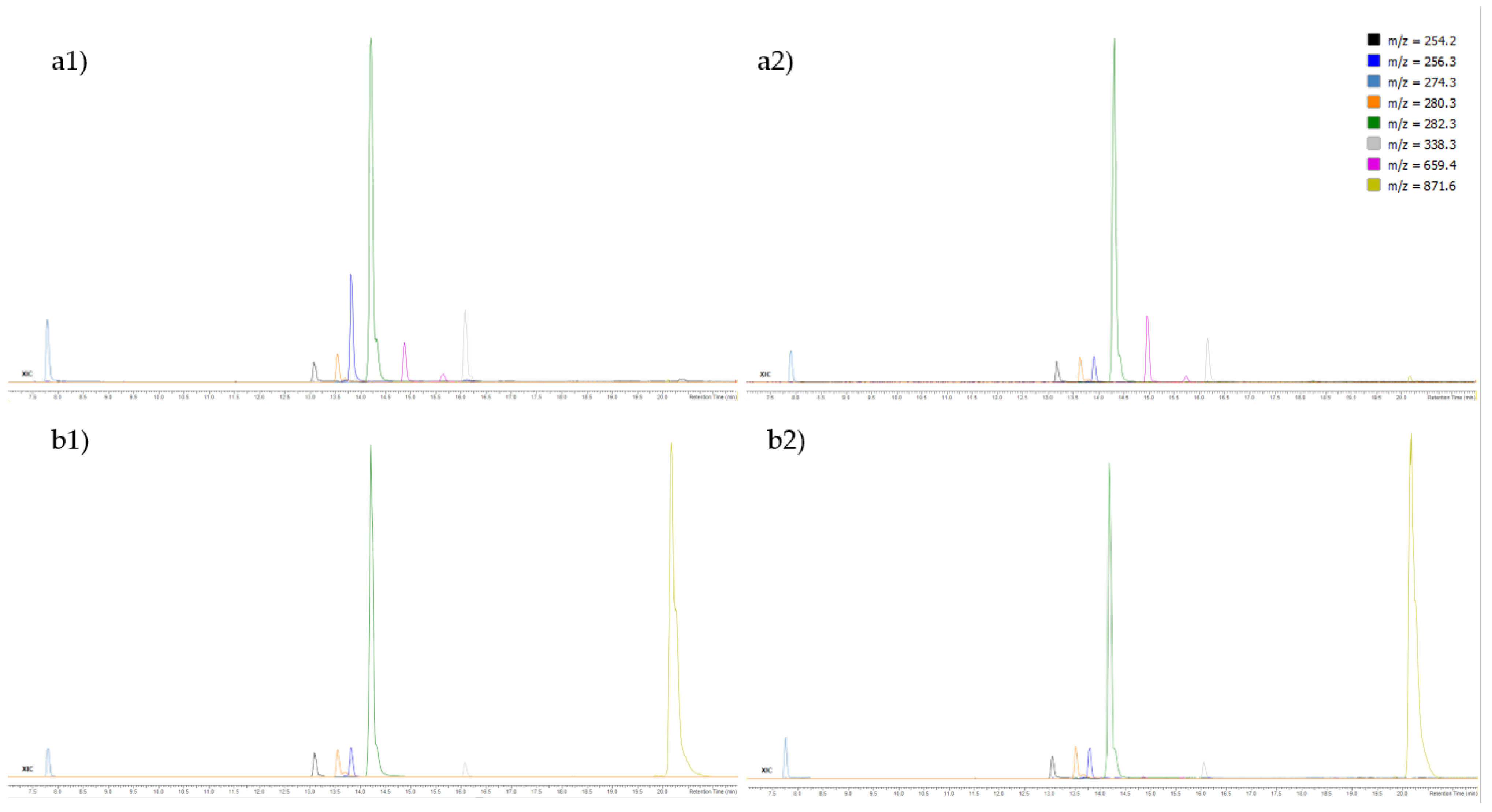
3.4. Ultra-High Performance Liquid Chromatography-High-Resolution Mass Spectrometry (UHPLC-ESI-HRMS) of F3 and F4 Fractions
3.5. Zebrafish Embryotoxicity Test
3.6. Antioxidant Activity of Tested Fractions
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bulleri, F.; Benedetti-Cecchi, L.; Acunto, S.; Cinelli, F.; Hawkins, S.J. The influence of canopy algae on vertical patterns of distribution of low-shore assemblages on rocky coasts in the northwest Mediterranean. J. Exp. Mar. Bio. Ecol. 2002, 267, 89–106. [Google Scholar] [CrossRef]
- Cheminée, A.; Sala, E.; Pastor, J.; Bodilis, P.; Thiriet, P.; Mangialajo, L.; Cottalorda, J.-M.; Francoura, P. Nursery value of Cystoseira forests for Mediterranean rocky reef fishes. J. Exp. Mar. Bio. Ecol. 2013, 442, 70–79. [Google Scholar] [CrossRef]
- United Nations Environment Programme (UNEP). Convention for the Protection of the Marine Environment and the Coastal Region of the Mediterranean and Its Protocols. Mediterranean Action Plan-Barcelona Convention Secretariat. 2019, p. 153. Available online: https://wedocs.unep.org/bitstream/handle/20.500.11822/31970/bcp2019_web_eng.pdf (accessed on 5 December 2021).
- Council of Europe. Bern Convention/Convention de Berne: Convention on the Conservation of European Wildlife and Natural Habitats/Convention Relative à la Conservation Dela Vie Sauvage et du Milieu Naturel de l’Europe; Appendix/Annexe I, 19.IX.1979; Council of Europe: Strasbourg, France, 1979. [Google Scholar]
- Directive, H. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union 1992, 206, 7–50. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: http://www.algaebase.org (accessed on 5 December 2021).
- Orellana, S.; Hernández, M.; Sansón, M. Diversity of Cystoseira sensu lato (Fucales, Phaeophyceae) in the eastern Atlantic and Mediterranean based on morphological and DNA evidence, including Carpodesmia gen. emend. and Treptacantha gen. emend. Eur. J. Phycol. 2019, 54, 447–465. [Google Scholar] [CrossRef]
- Molinari-Novoa, E.A.; Guiry, M.D. Reinstatement of the genera Gongolaria Boehmer and Ericaria Stackhouse (Sargassaceae, Phaeophyceae). Not. Algarum 2020, 171, 1–10. [Google Scholar]
- Milkova, T.; Talev, G.; Christov, R.; Dimitrova-Konaklieva, S.; Popov, S. Sterols and volatiles in Cystoseira barbata and Cystoseira crinita from the Black sea. Phytochemistry 1997, 45, 93–95. [Google Scholar] [CrossRef]
- Ivanova, V.; Stancheva, M.; Petrova, D. Fatty acid composition of black sea Ulva rigida and Cystoseira crinite. Bulg. J. Agric. Sci. 2013, 19, 42–47. [Google Scholar]
- Kamenarska, Z.; Yalçin, F.N.; Ersöz, T.; Çaliş, I.; Stefanov, K.; Popov, S. Chemical composition of Cystoseira crinita Bory from the eastern Mediterranean. Z. Naturforsch. 2002, 57, 584–590. [Google Scholar] [CrossRef]
- Fisch, K.M.; Böhm, V.; Wright, A.D.; König, G.M. Antioxidative meroterpenoids from the brown alga Cystoseira crinita. J. Nat. Prod. 2003, 66, 968–975. [Google Scholar] [CrossRef]
- Praud, A.; Valls, R.; Piovetti, L.; Banaigs, B.; Benaim, J.-Y. Meroditerpenes from the brown alga Cystoseira crinita off the French Mediterranean coast. Phytochemistry 1995, 40, 495–500. [Google Scholar] [CrossRef]
- Gouveia, V.; Seca, A.M.; Barreto, M.C.; Pinto, D.C. Di-and sesquiterpenoids from Cystoseira genus: Structure, intra-molecular transformations and biological activity. Mini Rev. Med. Chem. 2013, 13, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, C.B.; Gangadhar, K.N.; Macridachis, J.; Pavão, M.; Morais, T.R.; Campino, L.; Varela, J.; Lago, J.H.G. Cystoseira algae (Fucaceae): Update on their chemical entities and biological activities. Tetrahedron Asymmetry 2017, 28, 1486–1505. [Google Scholar] [CrossRef]
- Amico, V.; Cunsolo, F.; Piatteli, M.; Ruberto, G.; Mayol, L. Strictaketal, a new tetraprenyltoluquinol with a heterotetracyclic diterpene moiety from the brown alga Cystoseira stricta. J. Nat. Prod. 1987, 50, 449–454. [Google Scholar] [CrossRef]
- Amico, V.; Cunsolo, F.; Piatteli, M.; Ruberto, G. Prenylated O-methyltoluquinols from Cystoseira stricta. Phytochemistry 1987, 26, 1719–1722. [Google Scholar] [CrossRef]
- Amico, V.; Oriente, G.; Neri, P.; Piatteli, M.; Ruberto, G. Tetraprenyltoluquinols from the brown alga Cystoseira stricta. Phytochemistry 1987, 26, 1715–1718. [Google Scholar] [CrossRef]
- Amico, V.; Piatteli, M.; Cunsolo, F.; Neri, P.; Ruberto, G. Two epimeric, irregular diterpenoid toluquinols from the brown alga Cystoseira stricta. J. Nat. Prod. 1989, 52, 962–969. [Google Scholar] [CrossRef]
- Mesguiche, V.; Valls, R.; Piovetti, L.; Banaigs, B. Meroditerpenes from Cystoseira amentacea var. stricta collected off the mediterranean coasts. Phytochemistry 1997, 45, 1489–1494. [Google Scholar] [CrossRef]
- De La Fuente, G.; Fontana, M.; Asnaghi, V.; Chiantore, M.; Mirata, S.; Salis, A.; Damonte, G.; Scarfì, S. The Remarkable Antioxidant and anti-inflammatory potential of the extracts of the brown alga Cystoseira amentacea var. stricta. Mar. Drugs 2021, 19, 2. [Google Scholar] [CrossRef]
- Bouafif, C.; Messaoud, C.; Boussaid, M.; Langar, H. Fatty acid profile of Cystoseira C. Agardh (Phaeophyceae, Fucales) species from the Tunisian coast: Taxonomic and nutritional assessments. Cienc. Mar. 2018, 44, 169–183. [Google Scholar] [CrossRef] [Green Version]
- Cravatt, B.F.; Prospero-Garcia, O.; Siuzdak, G.; Gilula, N.B.; Henriksen, S.J.; Boger, D.L.; Lerner, R.A. Chemical characterization of a family of brain lipids that induce sleep. Science 1995, 268, 1506–1509. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.-S.; Lee, S.R.; Jeong, Y.L.; Park, D.W.; Cho, Y.M.; Joo, H.M.; Kim, I.; Seu, Y.B.; Sohn, E.H.; Kang, S.C. Antiallergic activity of ethanol extracts of Arctium lappa L. undried roots and its active compound, oleamide, in regulating FcεRI-mediated and MAPK signaling in RBL-2H3 cells. J. Agric. Food Chem. 2016, 64, 3564–3573. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Shkrob, I.; Rozentsvet, O.A. Fatty acid amides from freshwater green alga Rhizoclonium hieroglyphicum. Phytochem. 2000, 54, 965–967. [Google Scholar] [CrossRef]
- Bertin, M.J.; Zimba, P.V.; Beauchesne, K.R.; Huncik, K.M.; Moeller, P.D.R. Identification of toxic fatty acid amides isolated from the harmful alga Prymnesium parvum carter. Harmful Algae 2012, 20, 111–116. [Google Scholar] [CrossRef]
- Rueda-Orozco, P.E.; Montes-Rodriguez, C.J.; Ruiz-Contreras, A.E.; Mendez-Diaz, M.; Prospero-Garcia, O. The effects of anandamide and oleamide on cognition depend on diurnal variations. Brain Res. 2017, 1672, 129–136. [Google Scholar] [CrossRef]
- Langstein, J.; Hofstäder, F.; Schwarz, H. cis-9,10-Octadecenoamide, an endogenous sleep-inducing CNS compound, inhibits lymphocyte proliferation. Res. Immunol. 1996, 147, 389–396. [Google Scholar] [CrossRef]
- Ranger, C.M.; Winter, R.E.; Rottinghaus, G.E.; Backus, E.A.; Johnson, D.W. Mass spectral characterization of fatty acid amides from alfalfa trichomes and theirdeterrence against the potato leafhopper. Phytochemistry 2005, 66, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, W.; Matsui, K.; Akakabe, Y.; Itai, N.; Kajiwara, T. Volatiles from Zostera marina. Phytochemistry 1998, 47, 27–29. [Google Scholar] [CrossRef]
- Jerković, I.; Cikoš, A.-M.; Babić, S.; Čižmek, L.; Bojanić, K.; Aladić, K.; Ul’yanovskii, N.V.; Kosyakov, D.S.; Lebedev, A.T.; Čož-Rakovac, R.; et al. Bioprospecting of less-polar constituents from endemic brown macroalga Fucus virsoides J. Agardh from the Adriatic Sea and targeted antioxidant effects in vitro and in vivo (zebrafish model). Mar. Drugs 2021, 19, 235. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, A.; Naughton, L.M.; Dobson, A.D.W.; Rai, D.K. High-performance liquid chromatography/electrospray ionisation mass spectrometric characterisation of metabolites produced by Pseudovibrio sp. W64, a marine sponge derived bacterium isolated from Irish waters. Rapid Commun. Mass Spectrom. 2018, 32, 1737–1745. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.; Aoki, H.; Ramanathan, R.; Hait, N.C.; Takabe, K. Sphingosine-1-phosphate signaling in immune cells and inflammation: Roles and therapeutic potential. Mediat. Inflamm. 2016, 2016, 8606878. [Google Scholar] [CrossRef] [Green Version]
- Fischer, C.L.; Drake, D.; Dawson, D.V.; Blanchette, D.R.; Brogden, K.A.; Wertz, P.W. Antibacterial activity of sphingoid bases and fatty acids against gram-positive bacteria and gram-negative bacteria. Antimicrob. Agents Chemother. 2012, 56, 1157–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terasaki, M.; Hirose, A.; Narayan, B.; Baba, Y.; Kawagoe, C.; Yasui, H.; Saga, N.; Hosokawa, M.; Miyashita, K. Evaluation of recoverable functional lipid components of several brown seaweeds (Phaeophyta) from japan with special reference to fucoxanthin and fucosterol contents. J. Phycol. 2009, 45, 974–980. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Fukuda, S.; Izumi, H.; Saga, N. Anti-oxidant and fucoxanthin contents of brown alga ishimozuku (Sphaerotrichia divaricata) from the west coast of Aomori Japan. Mar. Drugs 2018, 16, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méresse, S.; Fodil, M.; Fleury, F.; Chénais, B. Fucoxanthin, a marine-derived carotenoid from brown seaweeds and microalgae: A promising bioactive compound for cancer therapy. Int. J. Mol. Sci. 2020, 21, 9273. [Google Scholar] [CrossRef] [PubMed]
- Osório, C.; Machado, S.; Peixoto, J.; Bessada, S.; Pimentel, F.B.; Alves, R.C.; Oliveira, M.B.P.P. Pigments content (chlorophylls, fucoxanthin and phycobiliproteins) of different commercial dried algae. Separations 2020, 7, 33. [Google Scholar] [CrossRef]
- Haugan, J.A.; Liaaen-Jensen, S. Algal carotenoids 54. Carotenoids of brown algae (Phaeophyceae). Biochem. Syst. Ecol. 1994, 22, 31–41. [Google Scholar] [CrossRef]
- Haugan, J.A.; Liaaen-Jensen, S. Improved isolation procedure for fucoxanthin. Phytochemistry 1989, 28, 2797–2798. [Google Scholar] [CrossRef]
- Lalegerie, F.; Gager, L.; Stiger-Pouvreau, V.; Connan, S. The stressful life of red and brown seaweeds on the temperate intertidal zone: Effect of abiotic and biotic parameters on the physiology of macroalgae and content variability of particular metabolites. In Advances in Botanical Research; Elsevier BV: Amsterdam, The Netherlands, 2020; pp. 247–287. [Google Scholar]
- Radman, S.; Cikoš, A.-M.; Flanjak, I.; Babić, S.; Čižmek, L.; Šubarić, D.; Čož-Rakovac, R.; Jokić, S.; Jerković, I. Less polar compounds and targeted antioxidant potential (in vitro and in vivo) of Codium adhaerens C. Agardh 1822. Pharmaceuticals 2021, 14, 944. [Google Scholar] [CrossRef]
- Kuczynska, P.; Jemiola-Rzeminska, M.; Strzalka, K. Photosynthetic pigments in diatoms. Mar. Drugs 2015, 13, 5847–5881. [Google Scholar] [CrossRef]
- Ina, A.; Hayashi, K.-I.; Nozaki, H.; Kamei, Y. Pheophytin a, a low molecular weight compound found in the marine brown alga Sargassum fulvellum, promotes the differentiation of PC12 cells. Int. J. Dev. Neurosci. 2007, 25, 63–68. [Google Scholar] [CrossRef]
- Okai, Y.; Higashi-Okai, K. Potent anti-inflammatory activity of pheophytin a derived from edible green alga, Enteromorpha prolifera (Sujiao-nori). Int. J. Immunopharmacol. 1997, 19, 355–358. [Google Scholar] [CrossRef]
- Okai, Y.; Ishizaka, S.; Higashi-Okai, K.; Yamashita, U. Detection of immunomodulating activities in an extract of Japanese edible seaweed Laminaria japonica (Makonbu). J. Sci. Food Agric. 1996, 72, 455–460. [Google Scholar] [CrossRef]
- Cahyana, A.H.; Shuto, Y.; Kinoshita, Y. Pyropheophytin a as an antioxidative substance from the marine alga Arame (Eisenis bicyclis). Biosci. Biotechnol. Biochem. 1992, 56, 1533–1535. [Google Scholar] [CrossRef]
- Nishibori, S.; Namiki, K. Antioxidative substances in the green fractions of the lipid of Aonori (Enteromorpha sp.). J. Home Econ. 1998, 39, 1173–1178. [Google Scholar] [CrossRef]
- Yang, X.; Kang, M.-C.; Lee, K.-W.; Kang, S.-M.; Lee, W.-W.; Jeon, Y.-J. Antioxidant activity and cell protective effect of loliolide isolated from Sargassum ringgoldianum subsp. Coreanum. Algae 2011, 26, 201–208. [Google Scholar] [CrossRef]
- Ochi, M.; Watanabe, M.; Miura, I.; Taniguchi, M.; Tokoroyama, T. Amijiol, isoamijiol, and 14-deoxyamijiol, three new diterpe- noids from the brown seaweed Dictyota linearis. Chem. Lett. 1980, 9, 1229–1232. [Google Scholar] [CrossRef]
- Hung, M.W.; Zhang, Z.J.; Li, S.; Lei, B.; Yuan, S.; Cui, G.Z.; Hoi, P.; Chain, K.; Lee, S.M.Y. From omics to drug metabolism and high content screen of natural product in zebrafish: A new model for discovery of neuroactive compound. Evid. Based Complementary Altern. Med. 2012, 2012, 605303. [Google Scholar] [CrossRef] [PubMed]
- **a, S.; Wang, K.; Wan, L.; Li, A.; Hu, Q.; Zhang, C. Production, characterization, and antioxidant activity of fucoxanthin from the marine diatom Odontella aurita. Mar. Drugs 2013, 11, 2667–2681. [Google Scholar] [CrossRef]
- Yoshida, Y.; Niki, E. Antioxidant effects of phytosterol and its components. J. Nutr. Sci. Vitaminol. 2003, 49, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Pangestuti, R.; Kim, S.-K. Biological activities and health benefit effects of natural pigments derived from marine algae. J. Funct. Foods 2011, 3, 255–266. [Google Scholar] [CrossRef]
- Utakod, N.; Laosripaiboon, W.; Chunhachart, O.; Issakul, K. The efficiency and the correlation between testing methods on antimicrobial and antioxidant activities of selected medicinal essential oils. Int. Food Res. J. 2017, 24, 2616–2624. [Google Scholar]
- Babić, S.; Čižmek, L.; Maršavelski, A.; Malev, O.; Pflieger, M.; Strunjak-Perović, I.; Topić Popović, N.; Čož-Rakovac, R.; Trebše, P. Utilization of the zebrafish model to unravel the harmful effects of biomass burning during Amazonian wildfires. Sci. Rep. 2021, 11, 2527. [Google Scholar] [CrossRef] [PubMed]
- Maes, J.; Verlooy, L.; Buenafe, O.E.; De Witte, P.A.; Esguerra, C.V.; Crawford, A.D. Evaluation of 14 organic solvents and carriers for screening applications in zebrafish embryos and larvae. PLoS ONE 2012, 7, e43850. [Google Scholar] [CrossRef]
- ISO. Water Quality—Determination of the Acute Lethal Toxicity of Substances to a Freshwater Fish (Brachydanio Rerio Hamilton-Buchanan (Teleostei, Cyprinidae)); Parts 1–3; International Organization for Standardization: Geneva, Switzerland, 1996. [Google Scholar]
- Council Directive 2010/63/EU on the protection of animals used for scientific purposes. Off. J. Eur. Union 2010, 276, 33–79. [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate fluorescence reader in 96-well format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Brand-Williams, W.; Cuvelier, M.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
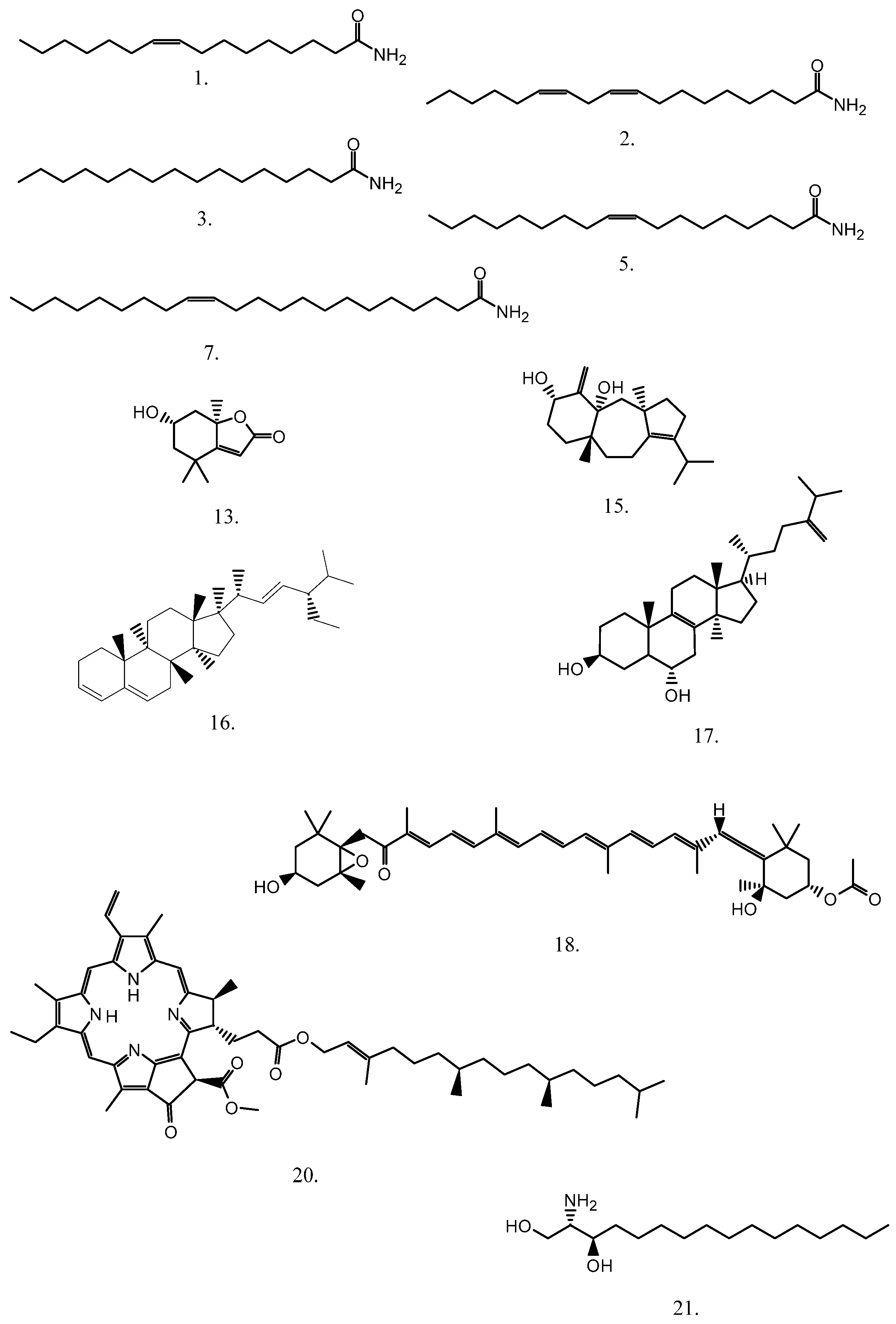
| No. | Compound | Elemental Composition | [M + H]+ | Error ** (ppm) | A | B | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| RT (min) | Area (Counts) | RT (min) | Area (Counts) | |||||||
| F3 | F4 | F3 | F4 | |||||||
| 1. | Palmitoleamide | C16H31NO | 254.24784 | 0.5 | 13.609 | 1108116 | 320801 | 13.085 | 1309202 | 3128998 |
| 2. | Linoleamide | C18H33NO | 280.26349 | 0.7 | 14.086 | 1390454 | 3166360 | 13.546 | 1574298 | 3629020 |
| 3. | Palmitamide | C16H33NO | 256.26349 | 0.9 | 14.460 | 5498614 | 3834048 | 13.818 | 6396427 | 3991527 |
| 4. | 2,3-Dihydroxypropyl palmitate | C19H38O4 | 331.28429 | 4.0 | 14.817 | 238983 | 16797 | 14.129 | 295785 | 21659 |
| 5. | Oleamide | C18H35NO | 282.27914 | 0.5 | 14.868 | 25284826 | 54228974 | 14.209 | 26096208 | 53855452 |
| 6. | 2,3-Dihydroxypropyl stearate | C21H42O4 | 359.31559 | 4.0 | 15.974 | 330917 | 58977 | 15.162 | 409784 | 70222 |
| 7. | Erucamide | C22H43NO | 338.34174 | 0.1 | 16.981 | 5433654 | 1438264 | 16.084 | 4682127 | 1767616 |
| 8. | 2-Hydroxypropyl stearate | C21H42O3 | 343.32067 | 1.5 | 17.269 | 38981 | 1214 | 16.254 | 46373 | 1025 |
| 9. | (2S)-1-Hydroxy-3-(tetradecanoyloxy)-2-propanyl (9Z)-9-octadecenoate | C35H66O5 | 567.49830 | 1.3 | 19.231 | 285866 | 10547 | 17.976 | 348859 | 14426 |
| 10. | 3-Phorbinepropanoic acid, 3,4-didehydro-9-ethenyl-14-ethyl-24,25-dihydro-21-(methoxycarbonyl)-4,8,13,18-tetramethyl-20-oxo-, (2E)-3,7,11,15-tetramethyl-2-hexadecen-1-yl ester | C55H72N4O5 | 869.55755 | 1.1 | 20.373 | 4307 | 1110897 | 19.066 | 5350 | 1005380 |
| 11. | Methyl (3R,10Z,14Z,20Z,22S,23S)-12-ethyl-3-hydroxy-13,18,22,27-tetramethyl-5-oxo-23-(3-oxo-3-{[(2E,7R,11R)-3,7,11,15-tetramethyl-2-hexadecen-1-yl]oxy}propyl)-17-vinyl-4-oxa-8,24,25,26-tetraazahexacyclo [19.2.1.16,9.111,14.116,19.02,7]heptacosa-1(24),2(7),6(27),8,10,12,14,16,18,20-decaene-3-carboxylate | C55H74N4O7 | 903.56303 | 1.8 | 20.441 | 45606 | 2940710 | 19.987 | 38274 | 2268701 |
| 12. | 3-Phorbinepropanoic acid, 9-acetyl-14-ethylidene-13,14-dihydro-21-(methoxycarbonyl)-4,8,13,18-tetramethyl-20-oxo-, 3,7,11,15-tetramethyl-2-hexadecen-1-yl ester | C55H74N4O6 | 887.56811 | 1.9 | 20.594 | 332532 | 11277103 | 19.953 | 401031 | 13903240 |
| 13. | Loliolide | C11H16O3 | 197.11722 | 1.2 | 6.225 | 88150 | 4603 | 6.323 | 86765 | 5693 |
| 14. | Isoamijiol oxidation product * | C20H30O2 | 303.23186 | 6.2 | 14.544 | 378362 | 2372 | 14.328 | 352017 | 2346 |
| 15. | (3aR,4aR,6S,8aR)-1-Isopropyl-3a,8a-dimethyl-5-methylene-2,3a,4,5,6,7,8,8a,9,10-decahydrobenzo[f]azulene-4a,6(3H)-diol (Isoamijiol) | C20H32O2 | 305.24751 | 0.5 | 15.974 | 122683 | 17514 | 15.522 | 104432 | 22138 |
| 16. | β-Stigmasterol | C29H46 | 395.36723 | 1.6 | 18.276 | 36308 | 364233 | 17.634 | 33268 | 459395 |
| 17. | (3β,6α)-14-Methylergosta-8,24(28)-diene-3,6-diol | C29H48O2 | 429.37271 | 3.4 | 18.395 | 784665 | 96119 | 17.480 | 980078 | 117567 |
| 18. | Fucoxanthin | C42H58O6 | 659.43062 | 2.1 | 14.873 | 2232648 | 3500 | 15.122 | 2697740 | 4095 |
| 19. | Pheophytin b | C55H72N4O6 | 885.55246 | 8.7 | 20.134 | 39490 | 467361 | 19.765 | 40024 | 391298 |
| 20. | Pheophytin a | C55H74N4O5 | 871.57320 | 2.1 | 20.815 | 5854 | 61538472 | 20.106 | 7418 | 57737924 |
| 21. | Hexadecasphinganine | C16H35NO2 | 274.27410 | 1.0 | 9.507 | 3186185 | 2813931 | 7.789 | 3952304 | 3361364 |
| No. | Compound | Elemental Composition | [M + H]+ | Error ** (ppm) | A | B | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| RT (min) | Area (Counts) | RT (min) | Area (Counts) | |||||||
| F3 | F4 | F3 | F4 | |||||||
| 1. | Palmitoleamide | C16H31NO | 254.24784 | 1.0 | 13.628 | 592811 | 2300496 | 13.05 | 497757 | 1965778 |
| 2. | Linoleamide | C18H33NO | 280.26349 | 2.0 | 14.088 | 479266 | 2697537 | 13.509 | 575290 | 2671906 |
| 3. | Palmitamide | C16H33NO | 256.26349 | 1.5 | 14.48 | 706476 | 3340508 | 13.781 | 628067 | 2844809 |
| 4. | 2,3-Dihydroxypropyl palmitate | C19H38O4 | 331.28429 | 3.1 | 14.82 | 18264 | 50976 | 14.105 | 17392 | 63085 |
| 5. | Oleamide | C18H35NO | 282.27914 | 1.5 | 14.888 | 4871406 | 36851920 | 14.189 | 6088715 | 30534284 |
| 6. | 2,3-Dihydroxypropyl stearate | C21H42O4 | 359.31559 | 1.5 | 15.994 | 21985 | 83888 | 15.14 | 19816 | 86090 |
| 7. | Erucamide | C22H43NO | 338.34174 | 0.6 | 16.994 | 2971406 | 1697387 | 16.057 | 2470961 | 1417294 |
| 8. | 2-Hydroxypropyl stearate | C21H42O3 | 343.32067 | 0.0 | 17.181 | 896 | 7775 | 16.327 | 1111 | 9880 |
| 9. | (2S)-1-Hydroxy-3-(tetradecanoyloxy)-2-propanyl (9Z)-9-octadecenoate | C35H66O5 | 567.49830 | 1.9 | 19.225 | 185815 | 176355 | 17.948 | 226104 | 218486 |
| 10. | 3-Phorbinepropanoic acid, 3,4-didehydro-9-ethenyl-14-ethyl-24,25-dihydro-21-(methoxycarbonyl)-4,8,13,18-tetramethyl-20-oxo-, (2E)-3,7,11,15-tetramethyl-2-hexadecen-1-yl ester | C55H72N4O5 | 869.55755 | 0.0 | 20.37 | 1042 | 1968855 | 20.01 | 866 | 1678772 |
| 11. | Methyl (3R,10Z,14Z,20Z,22S,23S)-12-ethyl-3-hydroxy-13,18,22,27-tetramethyl-5-oxo-23-(3-oxo-3-{[(2E,7R,11R)-3,7,11,15-tetramethyl-2-hexadecen-1-yl]oxy}propyl)-17-vinyl-4-oxa-8,24,25,26-tetraazahexacyclo [19.2.1.16,9.111,14.116,19.02,7]heptacosa-1(24),2(7),6(27),8,10,12,14,16,18,20-decaene-3-carboxylate | C55H74N4O7 | 903.56303 | 3.3 | 20.437 | 48785 | 1044658 | 19.981 | 40622 | 876040 |
| 12. | 3-Phorbinepropanoic acid, 9-acetyl-14-ethylidene-13,14-dihydro-21-(methoxycarbonyl)-4,8,13,18-tetramethyl-20-oxo-, 3,7,11,15-tetramethyl-2-hexadecen-1-yl ester | C55H74N4O6 | 887.56811 | 1.3 | 20.591 | 45728 | 1294640 | 19.964 | 37547 | 1053061 |
| 13. | Loliolide | C11H16O3 | 197.11722 | 5.6 | 6.222 | 32631 | 14304 | 6.403 | 27206 | 13071 |
| 14. | Isoamijiol oxidation product * | C20H30O2 | 303.23186 | 0.4 | 14.534 | 48292 | 8398 | 14.427 | 51830 | 10486 |
| 15. | (3aR,4aR,6S,8aR)-1-Isopropyl-3a,8a-dimethyl-5-methylene-2,3a,4,5,6,7,8,8a,9,10-decahydrobenzo[f]azulene-4a,6(3H)-diol (Isoamijiol) | C20H32O2 | 305.24751 | 5.3 | 15.959 | 22350 | 12061 | 15.58 | 27465 | 14464 |
| 16. | β-Stigmasterol | C29H46 | 395.36723 | 1.9 | 18.288 | 19126 | 670276 | 17.607 | 16495 | 562142 |
| 17. | (3β,6α)-14-Methylergosta-8,24(28)-diene-3,6-diol | C29H48O2 | 429.37271 | 2.2 | 18.39 | 171540 | 273113 | 17.47 | 143026 | 279999 |
| 18. | Fucoxanthin | C42H58O6 | 659.43062 | 0.4 | 15.125 | 2138127 | 89864 | 14.851 | 1811467 | 86705 |
| 19. | Pheophytin b | C55H72N4O6 | 885.55246 | 0.2 | 20.148 | 20692 | 155992 | 19.776 | 21058 | 131321 |
| 20. | Pheophytin a | C55H74N4O5 | 871.57320 | 0.9 | 20.829 | 240373 | 41982766 | 20.116 | 199581 | 46894376 |
| 21. | Hexadecasphinganine | C16H35NO2 | 274.27410 | 0.4 | 9.473 | 1226868 | 3731490 | 7.762 | 1053691 | 3149871 |
| Sample | IC50 Value, mg/mL | Confidence Interval (95%) | Hillslope |
|---|---|---|---|
| FdEc F3 | 1.177 | 1.049–1.346 | 2.58 |
| FdEc F4 | ND * | - | - |
| FdEa F3 | 0.072 | 0.067–0.077 | 2.477 |
| FdEa F4 | 1.060 | 0.986–1134 | 3.944 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radman, S.; Čižmek, L.; Babić, S.; Cikoš, A.-M.; Čož-Rakovac, R.; Jokić, S.; Jerković, I. Bioprospecting of Less-Polar Fractions of Ericaria crinita and Ericaria amentacea: Developmental Toxicity and Antioxidant Activity. Mar. Drugs 2022, 20, 57. https://doi.org/10.3390/md20010057
Radman S, Čižmek L, Babić S, Cikoš A-M, Čož-Rakovac R, Jokić S, Jerković I. Bioprospecting of Less-Polar Fractions of Ericaria crinita and Ericaria amentacea: Developmental Toxicity and Antioxidant Activity. Marine Drugs. 2022; 20(1):57. https://doi.org/10.3390/md20010057
Chicago/Turabian StyleRadman, Sanja, Lara Čižmek, Sanja Babić, Ana-Marija Cikoš, Rozelindra Čož-Rakovac, Stela Jokić, and Igor Jerković. 2022. "Bioprospecting of Less-Polar Fractions of Ericaria crinita and Ericaria amentacea: Developmental Toxicity and Antioxidant Activity" Marine Drugs 20, no. 1: 57. https://doi.org/10.3390/md20010057