1. Introduction
Forests play a key role in the global environment as hotspots of terrestrial biodiversity [
1,
2], sinks of atmospheric carbon [
3], and providers of multiple ecosystem services for human societies [
4,
5]. Green foliage is the main interface between a tree and its above-ground environment. It is involved in several key ecosystem processes, such as photosynthesis triggered by light interactions (i.e., absorption and scattering), as well as gas and water exchanges [
6]. To understand the process-based functional and structural forest changes is a long-term scientific objective [
7], and it can be supported by a multitude of optical remote sensing (RS) data and methods e.g., [
8,
9,
10,
11,
12,
13,
14].
Optical RS methods require good understanding of the interactions between incoming solar radiation and the vegetation media that are dominated by photosynthetically active foliage. Canopy structure has a particular influence on canopy reflectance characteristics, and especially in the near and short-wave infrared and microwave optical domains [
15,
16,
17]. Although light–canopy interactions can be studied effectively using physically based leaf and canopy radiative transfer models (RTMs), all RTMs rely on a certain level of simplification of vegetation canopy representations. Models simulating vegetation as a formation of horizontally homogeneous layers, such as the family of SAIL models by Verhoef [
18] and Verhoef and Bach [
19], the semi-discrete model of Gobron et al. [
20], and the flux decomposition model of Kallel [
21], are better suited to such structurally homogeneous canopies as those of crops, grasslands, or even tree crowns under the assumption of very high canopy closure and foliage density. Better suited to spatially heterogeneous and structurally complex forest canopies, however, are 3D RTMs working with tree architectures created from geometric shapes, such as the Flight model of North [
22], FRT model of Kuusk and Nilson [
23], or FLiES model of Kobayashi and Iwabuchi [
24]. Steadily increasing computational capabilities allow for radiative transfer simulations of highly complex 3D canopies wherein the level of detail reaches to the individual leaf or needle. Such examples include the Raytran model of Govaerts and Verstraete [
25], radiosity-graphics model of Qin and Gerstl [
26], scalable plant RT model of Bailey et al. [
27] and Discrete Anisotropic Radiative Transfer (DART) model of Gastellu-Etechgorry et al. [
28,
29].
In this study, we used the DART model, which simulates in-situ and RS acquisitions (LiDAR data and images in the optical and thermal spectral domains) and also the radiative budget of any landscape configuration, including terrain topography and atmosphere above the simulated scene [
29]. Currently, DART enables constructing representations of trees by (i) creating simple predefined geometric shapes internally, (ii) importing external user-defined 3D objects, or (iii) transforming the imported 3D objects into simplified geometrical shapes [
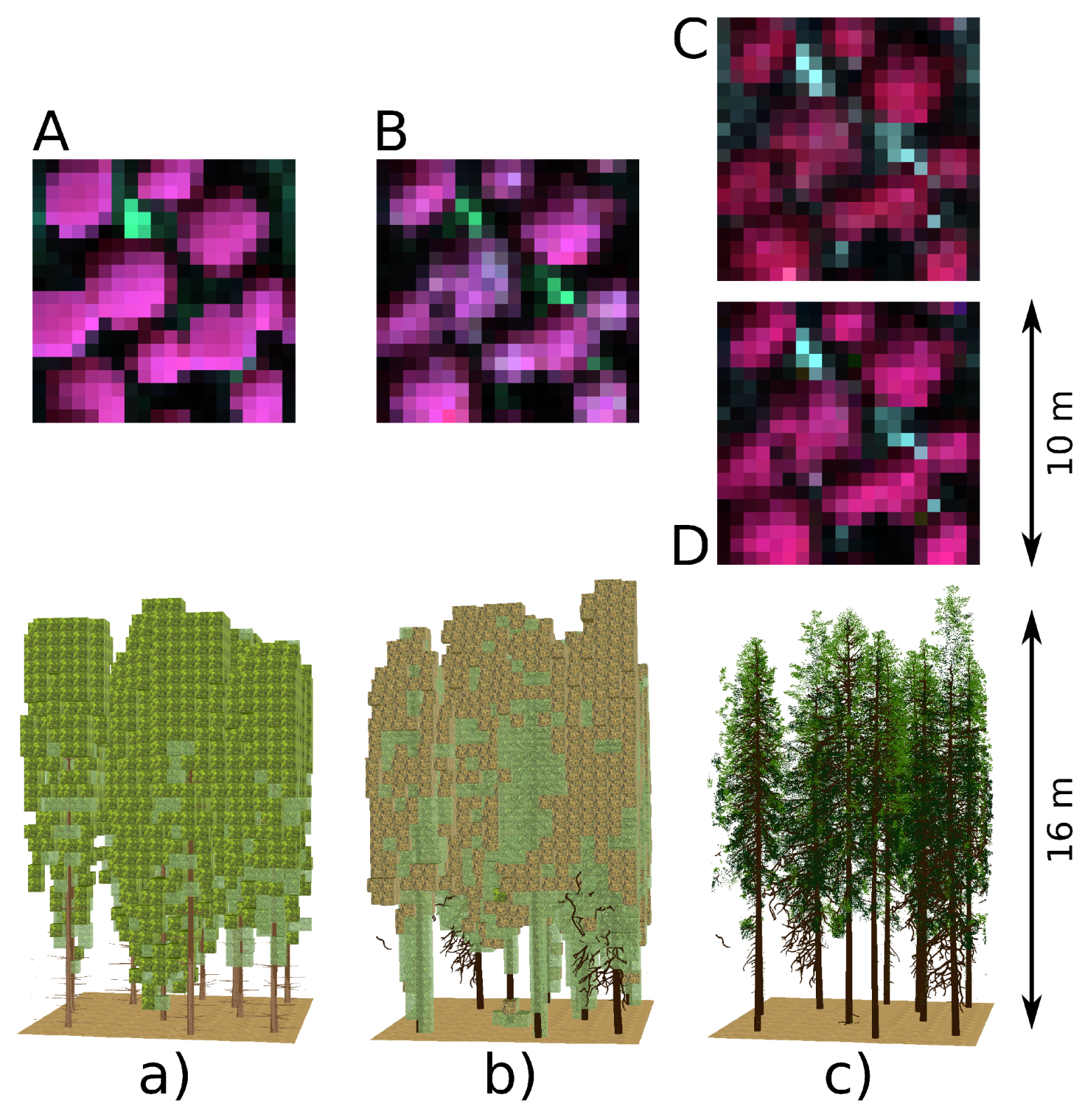
30]. The first of these utilities DART’s intrinsic functionalities defining a tree trunk and branches as simple 3D representations. Tree crown is represented by geometric shapes (e.g., an ellipsoid or a truncated cone) that are filled with triangles representing leaf facets or are created by cells known as voxels (i.e., 3D pixels) filled with a turbid medium representing foliage (
Figure 1a). The turbid medium represents an infinite number of infinitely small leaves. The triangular facets and the turbid medium are characterized by predefined leaf optical and structural properties. Structural properties are primarily controlled by leaf area index (LAI) and leaf angle distribution (LAD), both specified either per tree or per canopy of the entire simulated scene. Other controlling parameters are distribution of empty voxels (i.e., air gaps) and leaf volume density inside a voxel, which can be specified either per crown or according to crown vertical level. Such a tree representation is computationally very effective and well suited for forest canopies with relatively simple tree architecture, such as those of broadleaf trees. This parametrization may be less suitable for conifers, however, due to its insufficient description of fine-scale scattering properties of needle shoots representing the main foliage elements. Therefore, we hypothesize that the second manner of constructing tree representations, in this case from user-defined 3D objects (
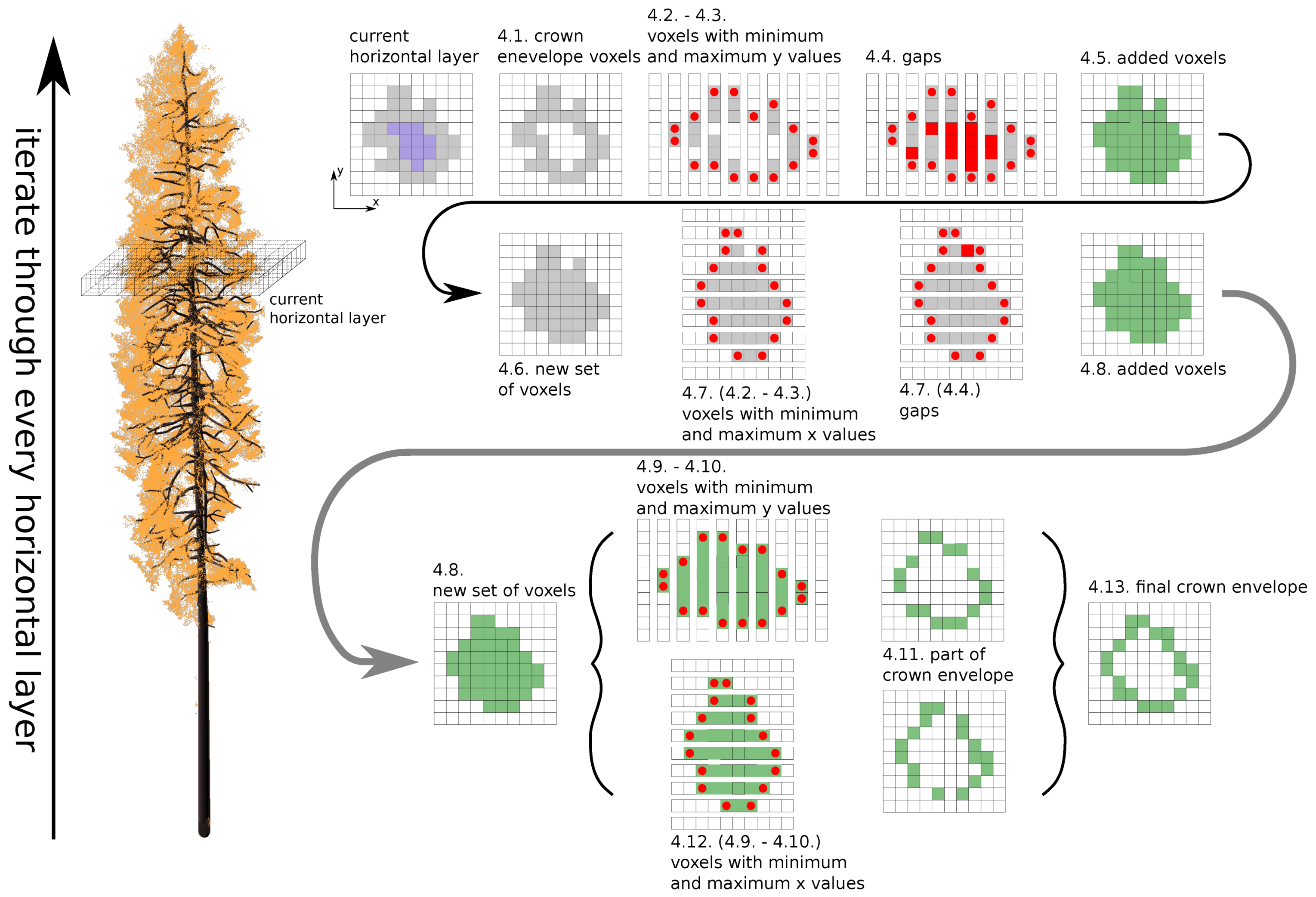
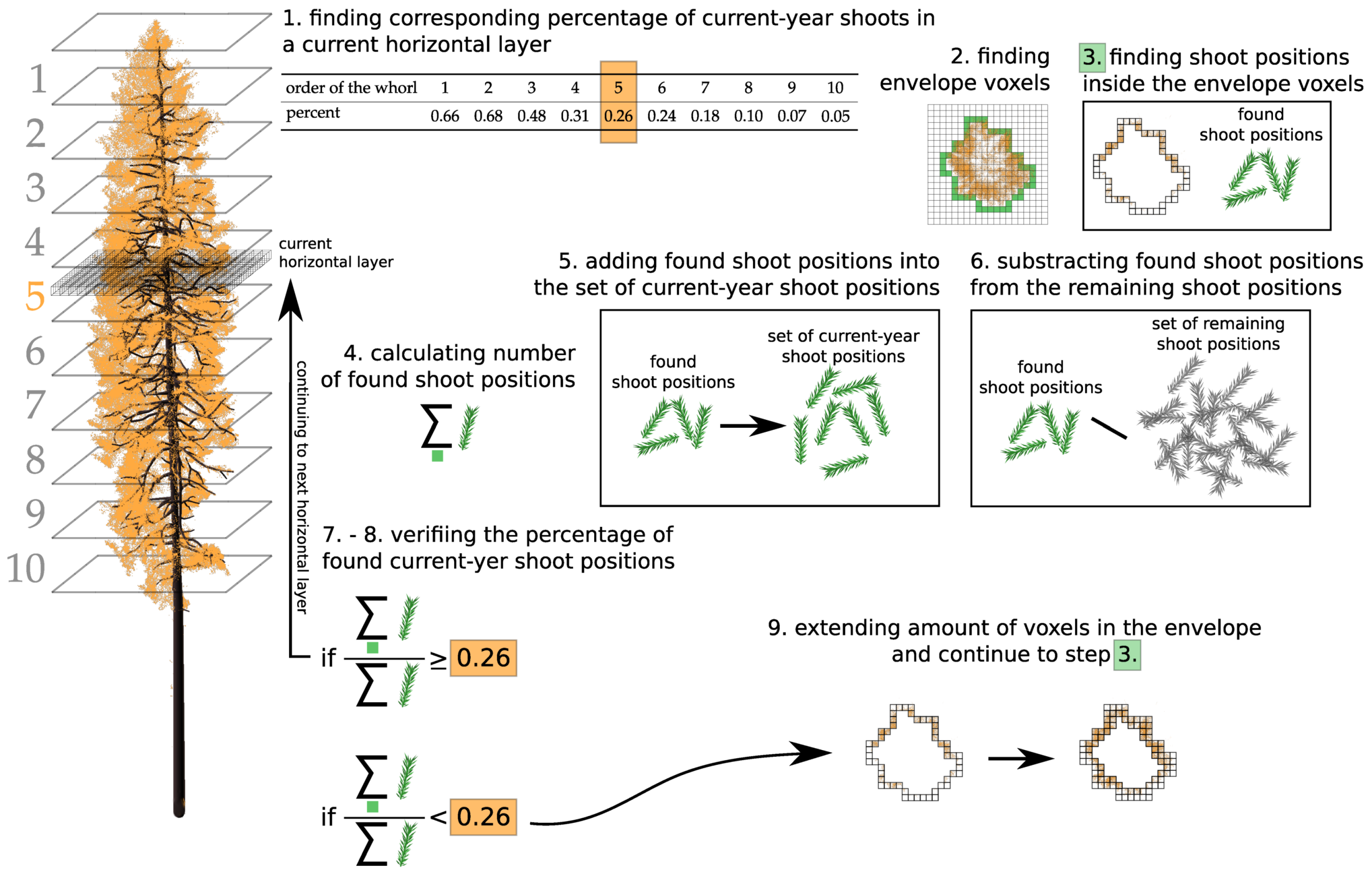
Figure 1b), might be more appropriate for conifers and allow for producing more accurate DART simulations. The approach imports trees as groups of geometrically explicit 3D objects with different optical properties (i.e., trunks, branches, needle shoots of different age classes, etc.). DART can be executed directly over the original 3D objects constructed from triangular facets, which represents structurally the most accurate solution. However, this option can be computationally highly demanding, especially when the imported 3D objects contains a lot of triangular facets. A third option is to transform the imported 3D foliage objects by voxelating them into leaf turbid cells (
Figure 1c). Although such transformed crown representations are structurally less precise, they preserve all original aspects of crown shapes (e.g., distribution of air gaps) and foliage distributions (i.e., LAI and LAD). The voxelated nature of a forest stand representation substantially accelerates the computation and subsequently makes DART more operational with regards to its inverse modelling.
Accurate physical representations that can account for complex 3D structures of ecosystems and link RS observations with studies of natural ecosystem processes are highly desirable. There is increasing demand, therefore, also for realistic 3D plant representations with natural architecture and foliage distribution. One group of methods for constructing 3D plants uses a set of parametric growing rules related to plant anatomy, topology, growth, and environmental factors derived from empirical observations. These are grammar-based methods, such as L-Systems [
31], Grogra [
32], OpenAlea [
33], and recently develo** computer graphic software (e.g., OnyxTree, Xfrog, and many others summarized at
http://vterrain.org/Plants/plantsw.html). For instance, Widlowski et al. [
34] used different virtual forests generated with Xforg and Arbaro software for the fourth phase of the radiative transfer model intercomparison (RAMI-IV) exercise. Nevertheless, most software tools create trees suitable only for exterior visualization and simulation of virtual landscapes (e.g., for computer games). They are less suitable for ecosystem studies requiring “biologically” correct 3D plant representations that account also for irregularities caused by natural and anthropogenic factors (e.g., spruce needle defoliation and regeneration; [
35]).
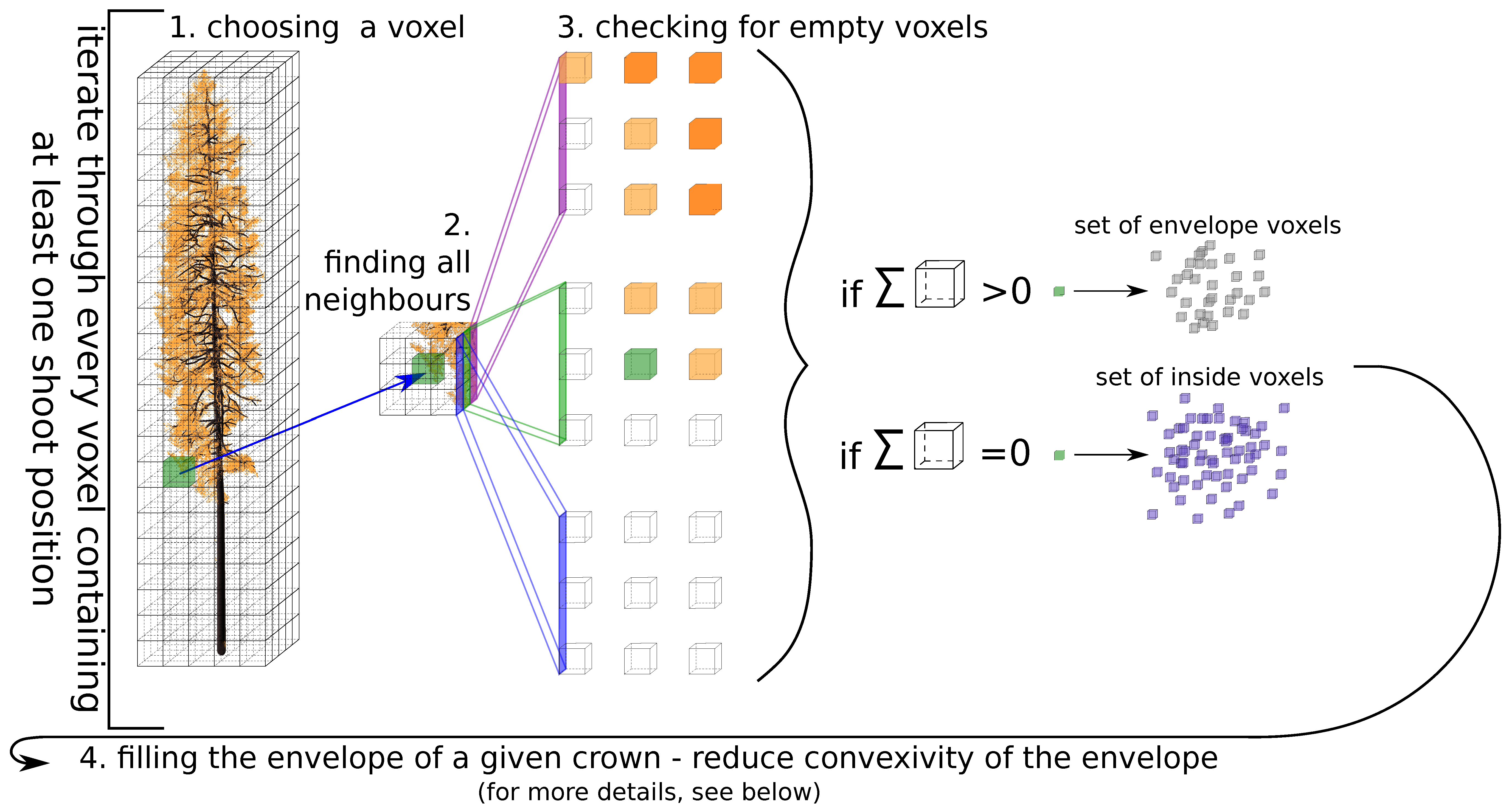
Another group of methods for constructing 3D tree representations uses the rapidly develo** technology of terrestrial laser scanning (TLS, also known as terrestrial LiDAR), which captures well the explicit 3D vegetation architecture [
36,
37,
38]. Several studies have reported reconstruction of tree wooden skeleton (stem and branches) using TLS data by constructing a quantitative structure model (QSM) of topologically and hierarchically ordered components [
39,
40,
41]. The TLS-based wooden skeleton reconstruction prefers data collected during leaf-off conditions, which means it is less than perfectly suited to conifers and evergreen-broadleaf mixed forests. In cases of evergreens, a TLS point cloud must be separated into wooden and foliage parts using (i) geometric properties [
42], (ii) radiometric properties [
43] or (iii) a combination of the two [
44].
Considerably fewer studies have reconstructed a whole tree by adding foliage elements to a wooden skeleton. These studies concern mainly broadleaf tree species [
45,
46,
47], and very few such attempts have been made for conifers [
48]. Statistical information about spatial density, size distribution, and angular orientation of flat leaves could be extracted from the point cloud data [
46,
49,
50]. For instance, Åkerblom et al. [
46] introduced an algorithm to insert primarily broadleaf geometric 3D representations into QSM, but its extension to complex needle leaf objects considerably increased the computation requirements. Côté et al. [
48] introduced the L-Architect model for conifers to reconstruct branching structure and to add foliage based on light availability derived from the point cloud data. Nonetheless, addressing 3D structure of conifers remains a challenge due to difficulties involving (i) leaf–wood separation, (ii) self-occlusion effects, and (iii) the complex geometrical nature of needle shoot elements.
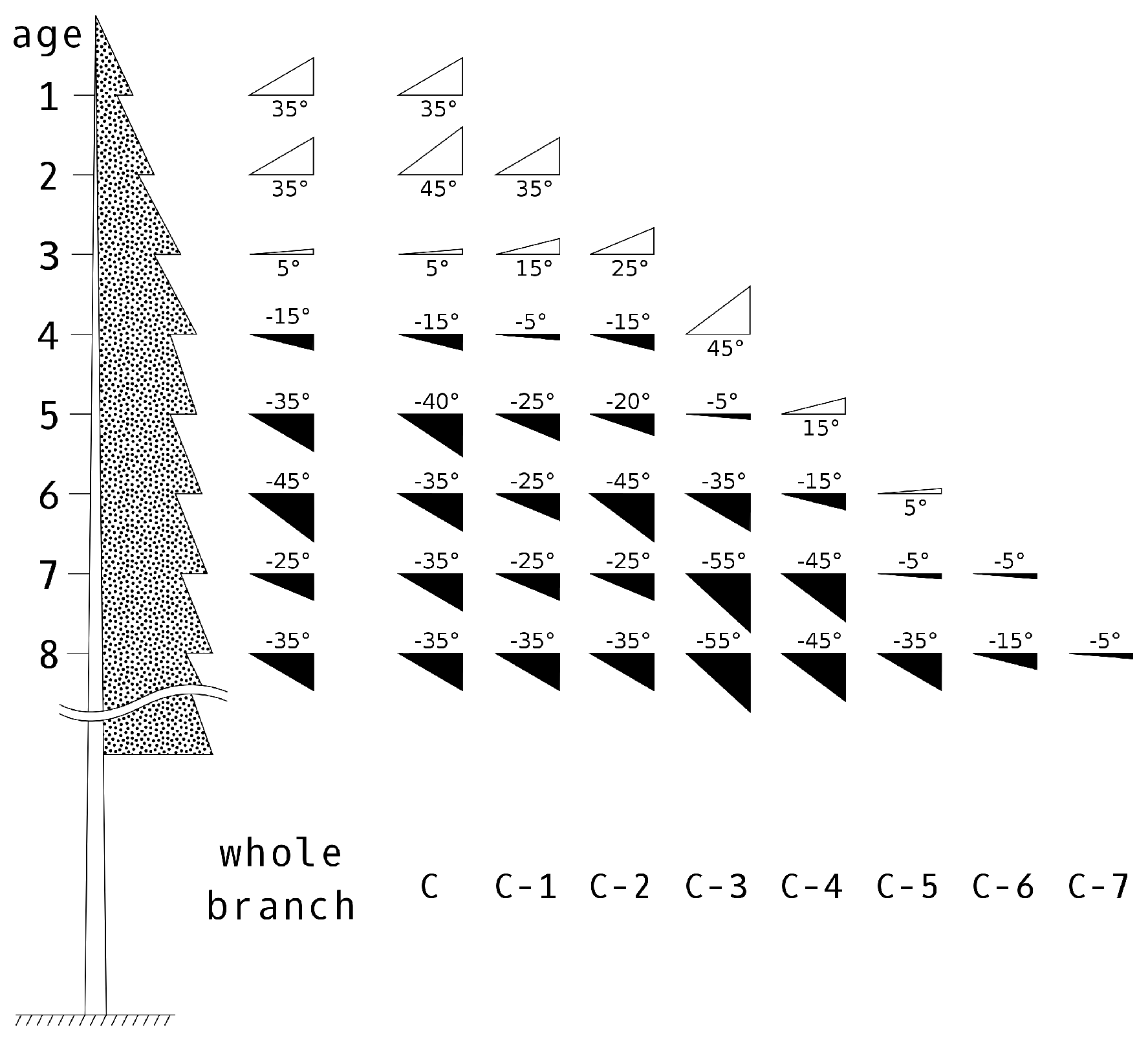
The objectives of this study, therefore, were to (i) design an algorithm for reconstructing a close-to-reality 3D representation of Norway spruce trees from TLS measurements (i.e., reconstruction of wooden skeleton and natural distribution of needle foliage) and (ii) evaluate the effect of canopy structure on forest reflectance as simulated in a radiative transfer model. The second objective was achieved through DART simulations of four scenarios using different levels of spruce canopy structural details. The results of the scenarios were cross-compared and assessed against corresponding airborne hyperspectral and Sentinel-2 MSI RS reflectance observations.
5. Conclusions
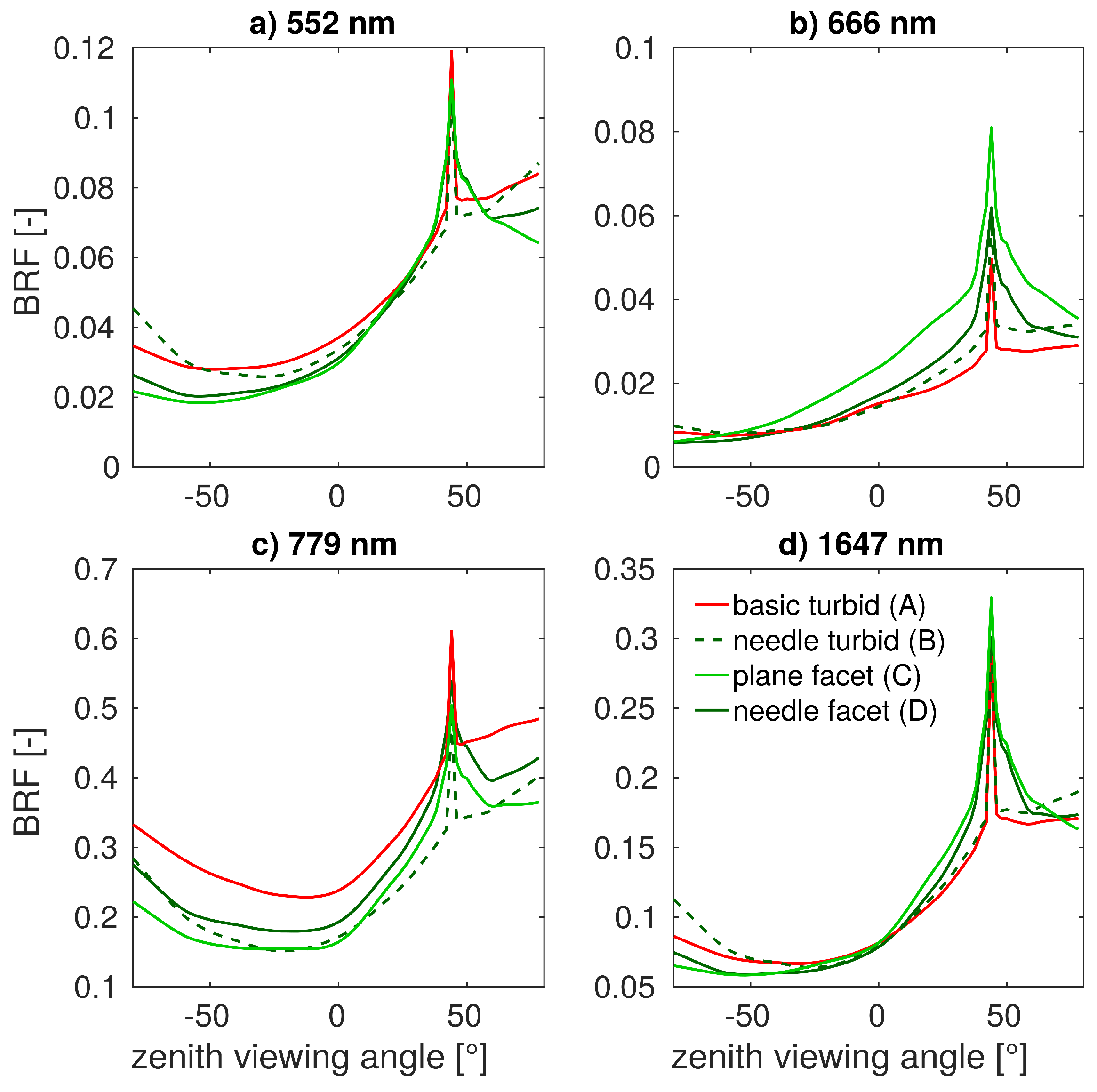
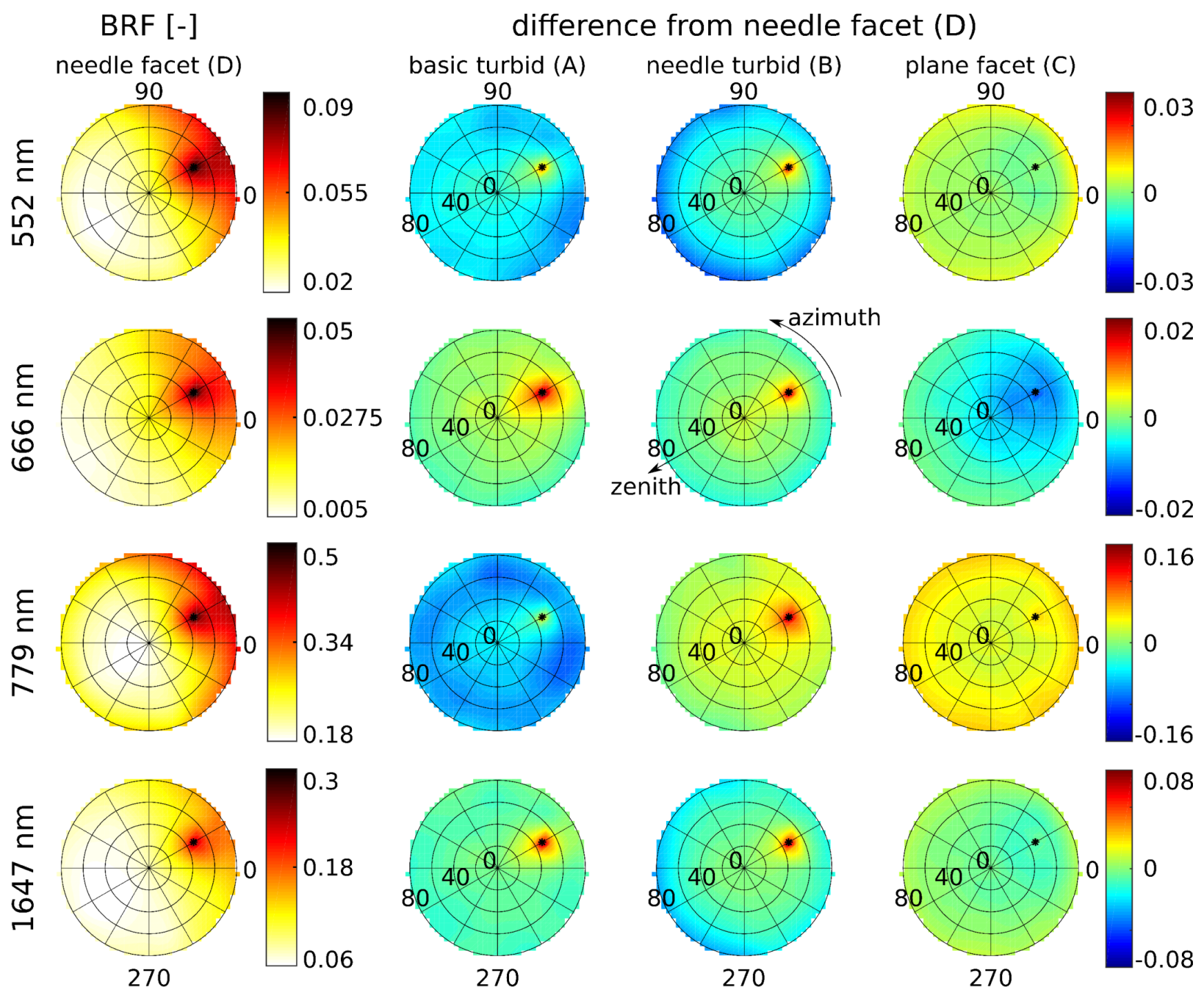
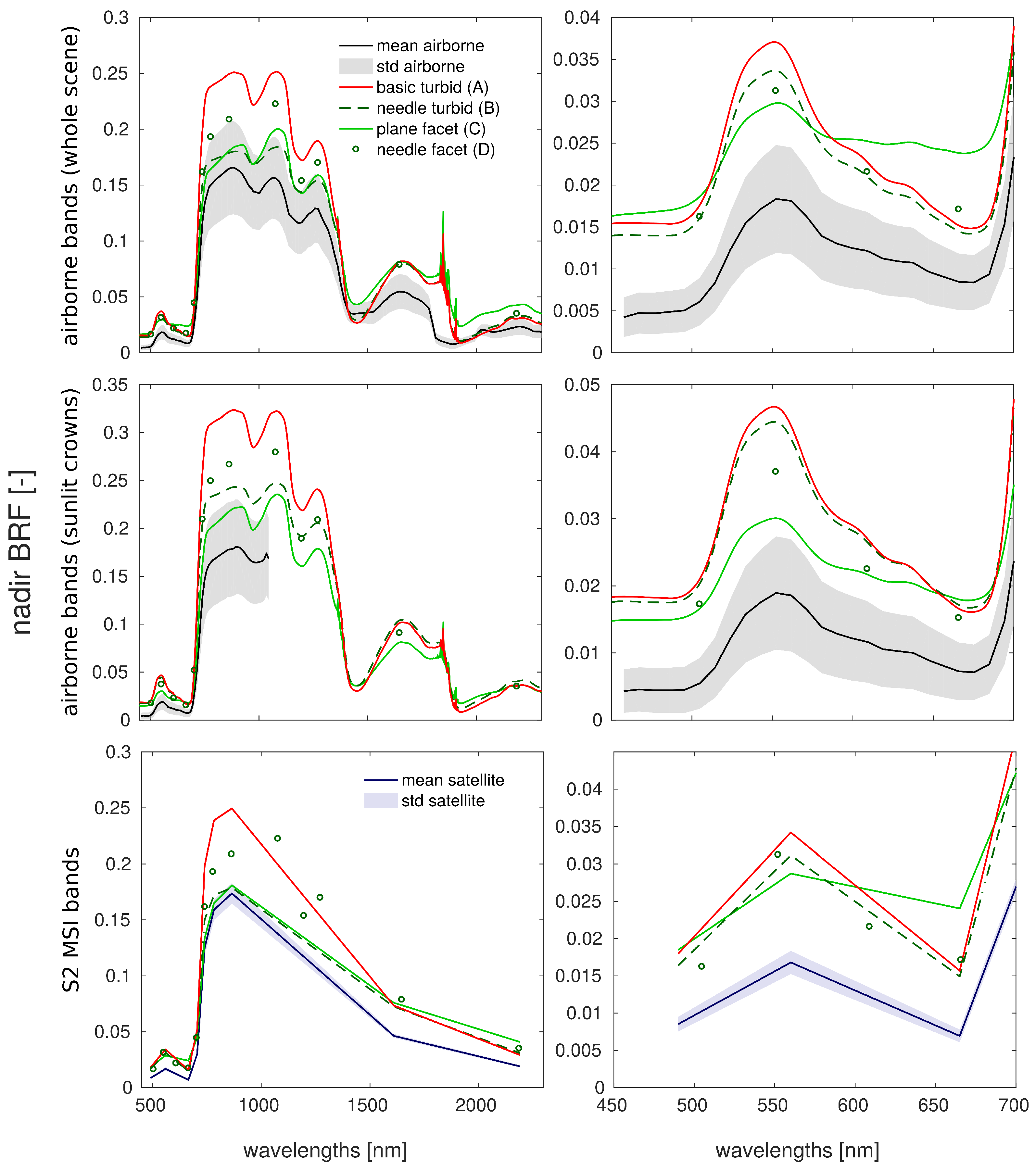
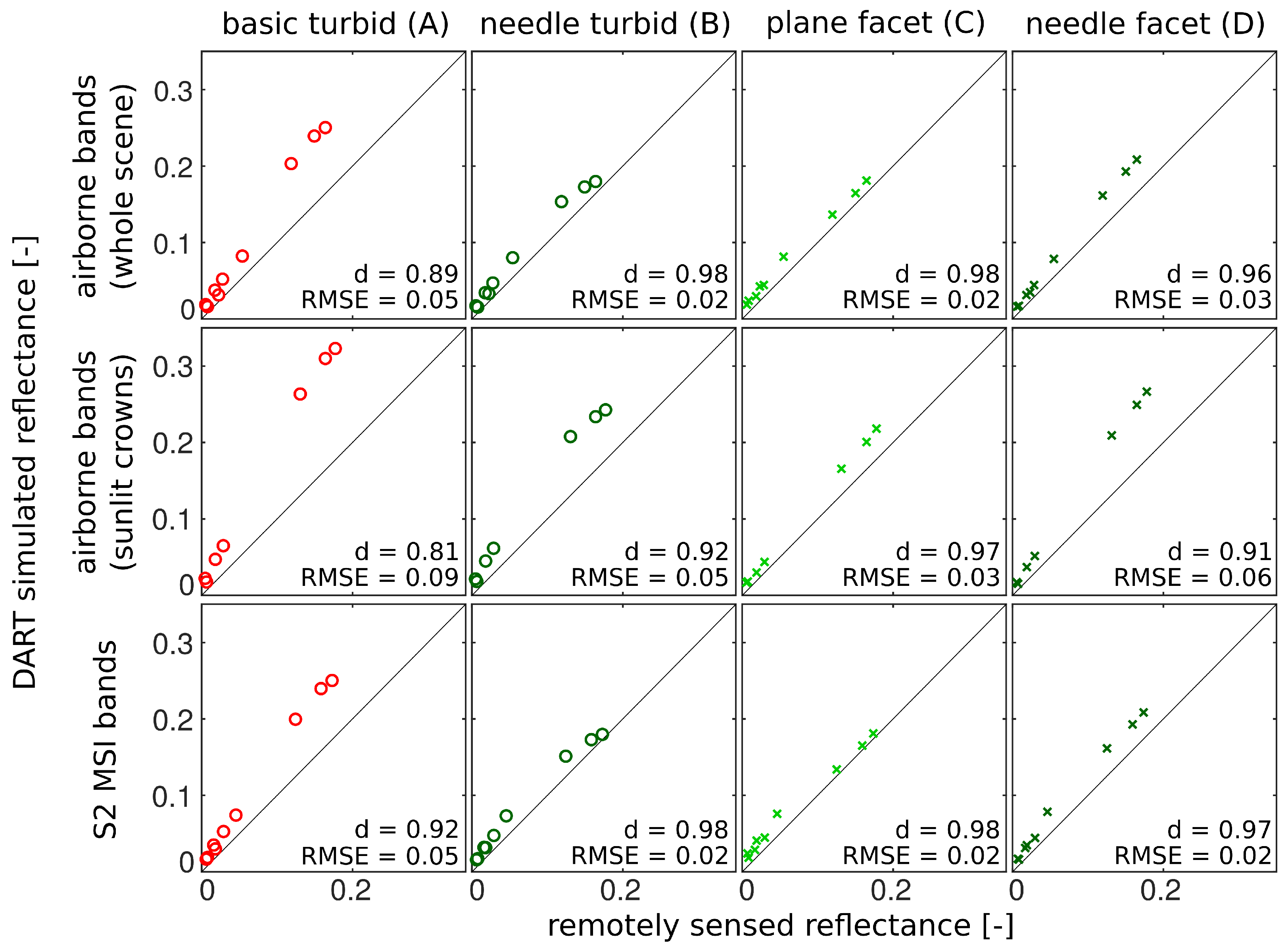
In this study, we evaluated the influence of Norway spruce forest structural representations on DART modelled top-of-canopy reflectance. DART simulated nadir reflectance values were compared with actual airborne hyperspectral and satellite multispectral images acquired over the simulated research site. Because the multi-angular directional reflectance measurements of the investigated spruce forest were not available, the multi-angular simulations were compared against the most detailed and presumably most accurate scenario D, needle facet. The results revealed improvements in reflectance accuracy simulated for scenarios wherein the 3D forest scene representations were retrieved and parametrized using the method designed in this study. We identified scenario B, needle turbid, as the most effective scenario for nadir simulations. This scenario was able to produce the visible and NIR reflectance with an acceptable level of accuracy (i.e., similar to that of scenario D and close to actual observations) with relatively short computational time and without a need for high-performance computer resources. Being based on the needle turbid representation of the stand, however, scenario B could not very well reproduce reflectance around the hot-spot and in the extreme zenith angles. In the case of the multi-angular directional reflectance, scenario C, plane facet, tracks closely the most detailed scenario D, needle facet, while maintaining a comparable (in NIR) or even better match (in visible wavelengths) with the reflectance of RS data acquired from the nadir.
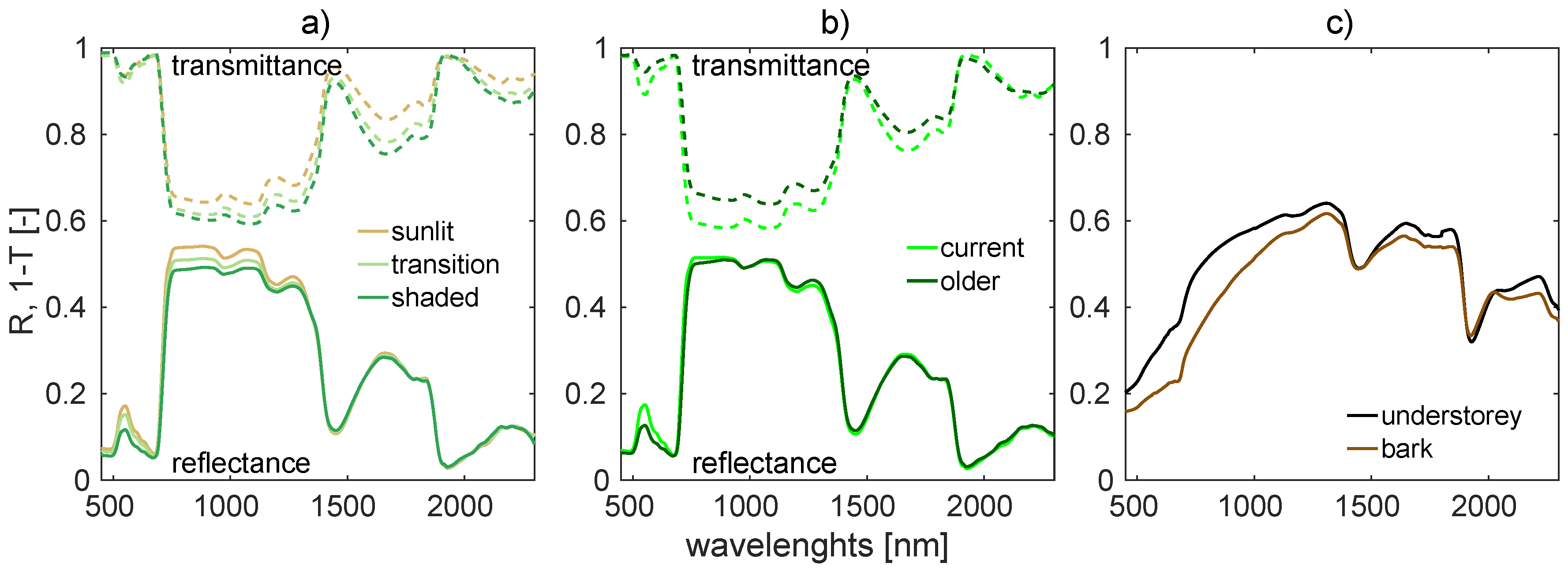
The 3D representation of Norway spruce trees reconstructed from TLS data with the semi-automatic algorithm designed in this study was found to reassemble adequately the habitus of real Norway spruce trees and increase the fidelity of canopy reflectance simulations in DART. The topographically accurate capturing of the tree structures and more detailed parametrization of foliage optical properties throughout a tree vertical profile allowed for more accurate solar radiation scattering within a simulated forest stand. Because the results obtained showed reflectance overestimations in visible and NIR spectral regions when compared with RS observations, there clearly is room for further improvements. Subsequent research should be focused upon (i) more realistic distribution of optical properties for wooden parts of forest stands (especially for young branches and tips of tree crowns), and (ii) the extension and improvement of needle shoot representations. Several needle shoot types could be reconstructed and used instead of a single, universal 3D model. Their size and geometrical parameters should reflect natural variability driven by shoot location within the tree crown, age, needle angular distribution, and within-shoot clum** in order to produce more realistic spruce tree representations for purposes of estimating radiative transfer.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}













