4.1. Literature and Database Searches (Epidemiological Studies)
Studies conducted prior to the SARS-CoV-2 pandemic era have already shown an association between the DD genotype and incidence, morbidity, and mortality risk in patients with acute respiratory distress syndrome (ARDS) [
39]. In addition, the frequency of the D allele was shown to be significantly higher in the hypoxemic group than in the non-hypoxemic group, whereas there was no significant difference between SARS cases and the control [
40].
According to our database analysis, the
ACE1 II genotype frequency in a population was significantly negatively correlated with the number of deaths due to SARS-CoV-2 infection, suggesting that the
ACE1 II genotype may favorably influence the prevalence and clinical outcome of COVID-19 [
1]. In addition, a detailed systematic review published by Pabalan et al. shows that the
ACE1 DD genotype may be an important prognostic marker for mortality in Asian COVID-19 patients with acute lung injury / ARDS [
41]. Similarly, by conducting an epidemiological study in the population from 26 Asian countries, Pati et al. showed that the
ACE1 D allele is associated with susceptibility to SARS-CoV-2 infection and with the mortality rate [
42]. These authors found a significant positive correlation of the D allele of
ACE1 polymorphism with SARS-CoV-2 infection and concluded that the D allele of
ACE1 I/D polymorphism is associated with the rate of infection and mortality. Two other groups have also shown that as the frequency of the
ACE1 II genotype increased, the mortality rate of COVID-19 decreased significantly [
43,
44].
Conversely, however, certain studies have shown that DD genotype is favorable in SARS-CoV-2 infection, or that there is no association between DD genotype frequency and COVID-19 mortality. Delanghe et al. conducted a multiple regression analysis using the data from 25 European countries and showed a negative correlation, i.e., the COVID-19 infection rate decreases as the frequency of the
ACE1 D allele increases [
45]. Meanwhile, Cenanovic et al. analyzed the
ACE1 D allele frequencies in 18 selected European populations and compared them with COVID-19 prevalence, mortality, and severity using multivariate linear regression analysis [
46]. Consequently, they found no clear statistical evidence to show that the
ACE1 D allele is associated with increased or decreased COVID-19 incidence, mortality, or case severity within the investigated populations.
4.2. Studies with Patient Samples (Clinical Studies)
Analysis using patient samples has been practiced in several countries on the Eurasian continent, providing valuable data (
Table 1). Calabrese et al. analyzed the association of severe/critical COVID-19 pneumonia with
ACE1 I/D polymorphism in southern Italy [
47]. Since the risk of thromboembolism is high, especially in patients with severe COVID-19 pneumonia, and decisive involvement of the balance between ACE1 and ACE2 activities is suggested in the thrombo-inflammatory process, 68 patients with severe COVID-19 pneumonia were divided according to the development of pulmonary embolism (PE) (25 PE + patients, 43 PE- patients). The results showed a statistically significant difference between PE + and PE- patients in assessing
ACE1 I/D polymorphisms (
p = 0.029). In particular, the prevalence of DD homozygous polymorphisms was significantly higher in PE + COVID-19 patients than in PE- patients (72 vs. 46.5%, respectively;
p = 0.048). Conversely, heterozygous I/D polymorphisms were significantly lower in PE + patients than in PE- patients.
ACE1 DD homozygosity has been associated with the development of thromboembolism in subjects without a predisposition to other diseases or changes in traditional thrombophilia. In this study, there were no significant differences in serum D-dimer levels between patients with COVID-19 with or without PE. However, the increased C-reactive protein observed in patients with PE suggests the important role of inflammation in the pathogenesis of thrombotic complications in patients affected by SARS-CoV-2 infection. In conclusion, the results of this study suggest that genetic factors are involved in the susceptibility to thromboembolism occurring in COVID-19.
ACE1 DD polymorphisms associated with high levels of both ACE1 and Ang II may represent genetic risk factors.
In southern Italy, a small study was also conducted using 27 patient samples, focusing on severe respiratory failure associated with COVID-19 pneumonia [
48]. At admission, 24 patients suffered from severe respiratory failure with paO2/FiO2 <100 mmHg and were treated with mechanical ventilation. Genetic testing of these patients showed that 19 (73%) of critically ill patients had DD genotype, whereas six (23%) had ID genotype. Only two patients (8%) showed II polymorphism. Studies on patients with ARDS prior to COVID-19 infection have demonstrated an association between DD genotype and cases, morbidity, and mortality risk.
In Spain, a comparative study was conducted using 204 age-matched controls with 204 COVID-19 patients (137 non-severe and 67 severe ICU cases) [
49]. Researchers determined the presence of
ACE1 I/D and
ACE2 rs2285666 polymorphisms and found that severe COVID-19 cases were associated with hypertensive male gender, hypertension, hypercholesterolemia, and the
ACE1 DD genotype (
Table 1 shows only the results of males). In multivariate logistic regression analysis, hypertension and male gender remained as independent and significant predictors of severity. The researchers also examined
ACE2 polymorphisms but found no association between them and disease outcomes. Poor outcomes of COVID-19 infection were associated with male gender, hypertension, hypercholesterolemia, and the
ACE1 genotype. The study suggested that
ACE1 I/D polymorphism may affect the severity of COVID-19, but the effect was dependent on the state of hypertension. The D allele was found to be significantly associated with hypoxemia in comparison to patients with non-hypoxemia. However, no association was found between individuals with the DD genotype and those with COVID-19 infection. In conclusion, the study states that the severity of COVID-19 symptoms may depend on age, diabetes, hypertension, and
ACE1 gene polymorphisms.
In India, Verma et al. genotyped
ACE1 I/D polymorphisms using RT-PCR with a sample of 269 COVID-19 patients and then performed a statistical analysis of the association between COVID-19 symptoms and
ACE1 I / D polymorphism [
50]. They found that the frequency of the
ACE1 DD genotype and D allele was significantly higher in patients with severe COVID-19.
ACE1 DD genotype, D allele frequency, older age (≥46 years), and presence of diabetes/hypertension were significantly higher in severe COVID-19 patient. The lack of an
ACE1 I allele was significantly correlated with patients with particularly severe COVID-19. These analyses were performed while adjusting for other factors such as age, gender, marriage status, income, diabetes, and hypertension, but the DD genotype was still significantly associated with clinical outcome of COVID-19, carrying a 3.6-fold higher risk.
A similar analytical study is being conducted for the first time in Turkey [
51]. Clinical data on COVID-19 in the Middle East are invaluable because if the D allele is indeed a risk factor, a big remaining puzzle is why the COVID-19 impact is much lower in the Middle East, where significantly more people have this allele as compared to the people of Europe. According to Gunal et al., age and the frequency of common comorbidities increased significantly in the infected group as the severity increased from asymptomatic as expected. The relationship between the
ACE1 genotype and severity was then analyzed, and they found that
ACE1 II genotype was the predominant genotype (50%) among asymptomatic patients, while the DD genotype was dominant among patients with severe symptoms (63.3%). This study indicates that the
ACE1 II genotype is protective against severe COVID-19.
Most of these findings suggest that heterozygous and non-II homozygous (DD + ID) genotypes of
ACE1 are risk factors for disease aggravation. Conversely, there is a controversial clinical report from the Czech Republic that does not match these data. Hubacek et al. analyzed 408 SARS-CoV-2-positive COVID-19 survivors (163 asymptomatic and 245 symptomatic) using Czech patient samples [
52]. The results showed that the frequency of
ACE1 II homozygotes was significantly higher in COVID-19 patients than in the controls. Importantly, this difference was only seen in symptomatic subjects. Thus, they concluded that
ACE1 I/D polymorphism could have the potential to predict the severity of COVID-19, with II homozygotes indicating an increased risk of symptomatic COVID-19. This result appears to be consistent with the epidemiological data published by Delanghe et al. showing an inverse correlation between the prevalence of COVID-19 infection and the frequency of the
ACE1 D allele [
45].
As described above, among the results using clinical samples, only Hubacek et al. showed the opposite results that II homozygotes, instead of ACE1 DD, may be associated with an increased risk of symptomatic COVID-19. The reason for these discrepancies is unknown at this time. They added the phrase "at least in Eastern European Caucasians" with that in mind and suggested that differences in geographical and ethnic factors may be behind it. Probably what is important in their study is the fact that Czech patients with genotype II were poor symptomatically, or at least there was no evidence that they were in better condition than those with the DD genotype. Since they analyzed only survivors, it is important to find out what the results would be if the deceased were included.
Meanwhile, we encountered an interesting article from Lebanon. Using 266 subjects (142 cases and 124 controls), the study found that individuals with the II genotype have a higher risk (OR = 2.373) of contracting COVID-19 as compared to the controls [
53]. These results seem to confirm Delanghe’s simulations in patients. Subjects with the DD genotype had a higher probability of experiencing severe COVID-19 symptoms conversely (OR = 7.173), to be hospitalized, and/or to be hypoxic. As to the reason the D allele becomes more resistant to infection, Delanghe et al. proposed a possible association of the D allele with a reduced expression of ACE2 [
45]. If this is the case, it will be supported from the virological point of view. However, this can also be a double-edged sword that reduces the beneficial effects of ACE2 on the lungs and other organs. Therefore, the loss of ACE2 can ultimately result in a detrimental enhancement of the Ang II/AT1R response. It will be interesting to investigate the possibility that DD genotype individuals are more resistant to viral infection initially but that the symptoms become more severe once infected as compared to those with
ACE1 II genotype. However, this theory does not well explain the low SARS-CoV-2 infection rate in East Asia, where the frequency of II is much higher than in other areas, such as Europe and the Middle East [
1,
54].
As to the deleterious effect of
ACE1 I/D polymorphism on COVID-19 symptoms, this situation seems to resemble the relationship between the
ACE1 DD type (D allele) and various comorbidities such as cardiovascular diseases [
55]. With regard to comorbidities, most of the database and clinical studies seem to support the involvement of the
ACE1 D allele and DD genotype, while some reports do not support it, and others deny it. However, this result is not surprising given that both comorbidities and COVID-19 are complicated diseases involving many factors in their establishment. In addition to further analysis in different regions and more patient samples, new analyses are expected that consider the involvement of other genes, as described below.
4.3. Possible Involvement of Polymorphic Alu Elements and Their Functional Aspects
Most of the genome is occupied by introns, and the gene exons that encode proteins make up only about 1% of our DNA. The interspersed repeats of mobility in our genome amount to about 45%, but they may be presumed to be non-functional "junk DNA", with the rare exceptions that cause genetic disease. This indicates that only a small part of the function of the entire genome is understood [
56]. Structural variations caused by Alu insertion have already been shown to be associated with the risk of many human diseases. The first known disease due to the insertion of a moving element was the insertion of a long scattered element-1 (LINE-1) that interrupted the coding exons of coagulation factor VIII, which is responsible for hemophilia A [
57]. Thus far, more than 120 LINE-1 mediated insertions causing hereditary diseases have been reported in humans [
58].
A growing body of evidence that Alus play an important regulatory role in gene expression in a wide range of physiological processes is accumulating [
59]. Recently, based on the hypothesis that a subset of common transposable polymorphisms affects human health, Payer et al. performed genome-wide association studies (GWAS) to identify a number of candidate loci for mutants caused by polymorphic Alu elements. They found 44 Alu insertion polymorphisms showing strong linkage disequilibrium (LD), and single nucleotide polymorphisms (SNPs) were most strongly associated with the disease phenotype [
58]. Therefore, it is likely that there is genetic evidence that the Alu mutant is functionally effective at these loci. Given that each insertion produces a structural feature of about 300 bp, the Alu variant is likely to result in causing genetic functional alterations, not just as a bystander.
Among many Alu elements, the
ACE1 polymorphism is a good example of how much research has been done thus far. Despite the fact that the Alu is an intron insertion, it can affect gene expression through several mechanisms, including both genetic and epigenetic pathways [
60]. At the molecular level, it has also been reported that the presence of the Alu element within intron 16 probably affects the promoter activity of
ACE1, which acts as a transacting repressor for RNA polymerase II activity [
61,
62]. Strikingly, ACE1 levels in the blood are determined by this I/D polymorphism [
63]. Individuals with the DD genotype have been shown to have significantly higher plasma ACE concentrations and ACE activity than people with type II and ID, showing complete LD with Alu DD.
4.4. Possible Interplay between ACE1 and Other Alu Variants
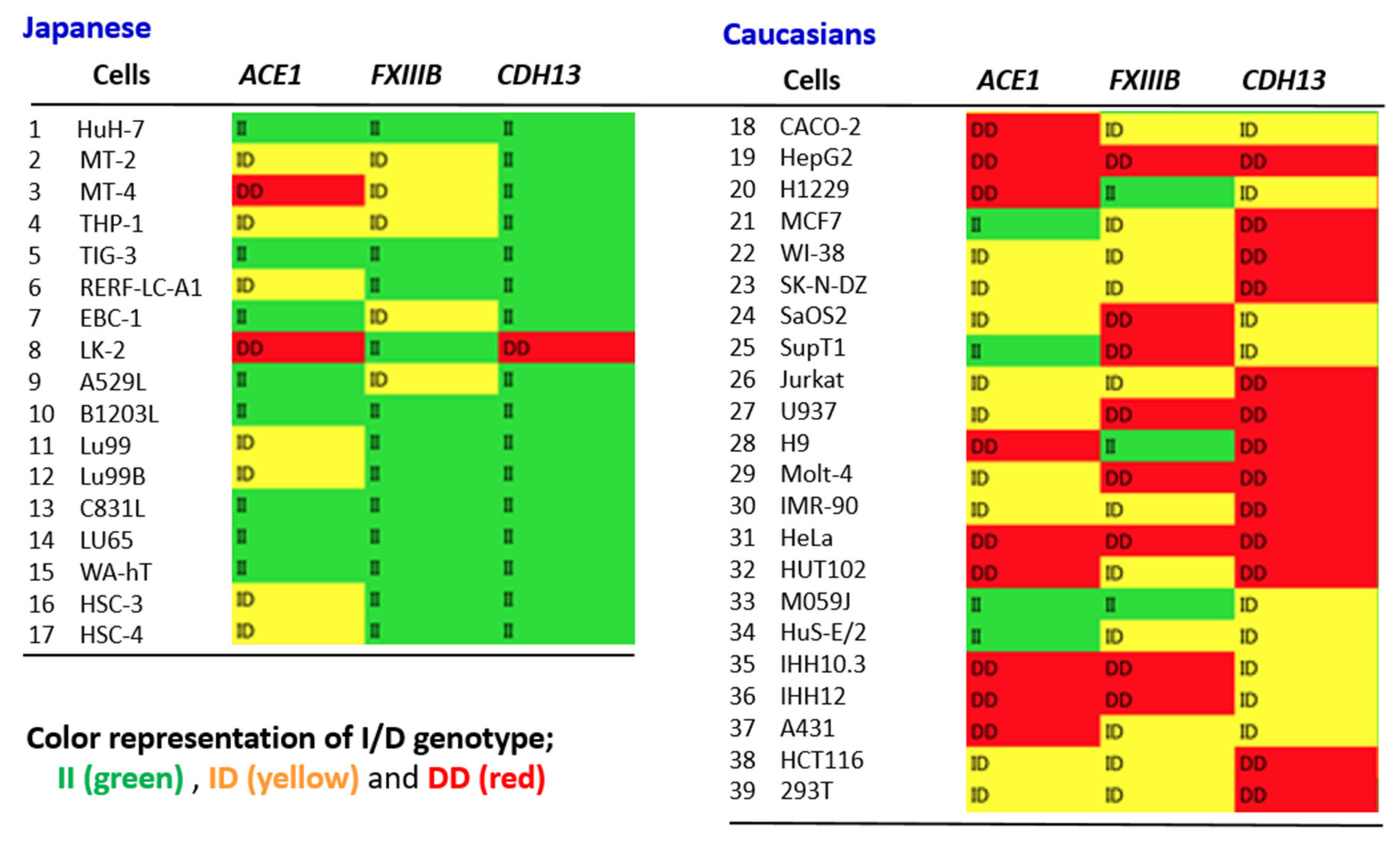
In searching for candidate genes that may act in cooperation with the
ACE1 variants, we have chosen the following criteria: the gene in question (1) has a function that can explain a systemic infection of COVID-19 that includes many organs and systems, (2) is associated with a comorbidity that leads to COVID-19 aggravation, and (3) shows an apparent ethnic difference in its genotype frequency between western and eastern populations. For this, we were particularly interested in the report by Wang et al. [
64]. They analyzed the frequency of
ACE1 Alu insertion and
APOBEC3 (A3B) deletion using a database. A3B proteins are editing enzymes that can interfere with the retrotransposition of endogenous retroelements such as the LINE and Alu. Surprisingly, Wang et al. found the variation curves for
ACE1 insertion and
A3B deletion among the geographic regions to be almost parallel, both rising continuously along the out-of-Africa expansion route. Alu retrotransposons have undergone many anthropological studies in the context of human evolution. Accordingly, these authors proposed an interesting hypothesis that functional loss of
A3B provided an opportunity for enhanced human adaptability and survival in response to the environmental and climate challenges arising during the migration from Africa. In this study, they also found that Alu insertion of other genes such as
FXIII and
CDH13 were increased, and these variants clearly showed ethnic differences in their genotypes [
64]. Human A3Bs can also interfere with the replication of exogenous retroviruses such as human immunodeficiency virus (HIV) and hepatitis B virus (HBV) [
65,
66]. Because intronic Alu is known to regulate gene expression through several mechanisms, both genetically and epigenetically, as described [
60], we then focused on the I/D polymorphisms of the
FXIIIB and
CDH13 genes, expecting the possibility of their interplay with
ACE1 variants.
FXIII is a transglutaminase and supports platelet adhesion and spreading as well as clot retraction, suggesting that FXIII is important for the stabilization of platelet-fibrin clots. FXIIIB has been studied in recent years for its association with the development of various hemorrhagic disorders, including ischemic stroke, venous thrombosis, coronary artery disease, and myocardial infarction [
67,
68,
69]. Importantly, FXIIIB functions antagonistically with tissue-type plasminogen activator (tPA) in the blood coagulation cascade. Recently, Matsuyama et al. proposed a possible role of the STAT3-PAI-1 signaling node in a chaotic chain reaction underlying COVID-19 pathophysiology. Viral proteins Nsp1 and Orf6, produced upon SARS-CoV-2 infection, efficiently inhibit STAT1 function in the cells. Suppressed STAT1 increases STAT3 activity, and STAT3 eventually upregulates PAI-1. This increased PAI-1 activity may result in IL-6 production through Toll-like receptor 4, which in turn activates more STAT3. Elevated STAT3 also activates PD-L1 in endothelial cells, leading to T-cell lymphopenia [
70].
FXIIIB polymorphism AluYa5 insertion has been reported to confer a risk of coronary atherosclerosis [
61]. Furthermore, intra- and extracellular FXIIIs are indicated to support the immobilization and killing of bacteria as well as phagocytosis by macrophages, strongly suggesting that FXIII may also function in innate immunity [
71]. With an analogy to the
ACE1 Alu I/D polymorphism, the
FXIIIB I/D type can also be involved in the difference in the susceptibility of Westerners and Asians against COVID-19, given the apparent difference in the genotype frequency seen between the those in the west and east.
Genetic variation of the cadherin gene
CDH13 was significantly correlated with certain clinical parameters, such as diastolic blood pressure, triglyceride, adiponectin, and insulin levels [
72]. The
CDH13 polymorphism is also associated with a decreased risk of develo** hypertension when compared to non-hypertensive individuals [
73]. A reduction in plasma T-cadherin levels is associated with increasing severity of coronary artery disease and a higher risk of acute coronary syndrome [
74]. Alu insertions of this gene, along with
ACE1,
FXIIIB, etc., are often studied to provide valuable information for ancestrality and genetic differentiation [
64]. However, the relationship between CDH13 and COVID-19 symptoms is not sufficiently studied at present.
Li et al. also investigated known Alu polymorphisms of genes such as
progesterone receptor (PGR) and
plasminogen activator (PLAT) in addition to the
ACE1 and
FXIIIB genes for their biological function, role in disease, and effect on COVID-19 [
60]. Of particular interest are the variations and functional changes in the
PGR gene, since this gene has already been shown to have reduced transcript and protein stability and reduced receptor responsiveness to progestin due to Alu intron insertion [
75]. This functional relationship of Alu with intron insertion is similar to the results seen with that of ACE1. However, the frequency of Alu insertion of the
PGR gene is significantly higher in Europeans than in the Asian population, which is the opposite of Alu polymorphisms in
ACE1,
FXIIIB, and
PLAT. Therefore, this suggests that the A3B rules do not apply to all Alu polymorphisms. Nevertheless, the effect of COVID-19 on clinical symptoms by Alu’s variant, which includes
PGR in addition to
ACE1 and
FXIIIB, may be worth further study. There is also a significant interaction between
PAI gene polymorphisms,
PAI4G5G and
ACE1 I/D polymorphisms, at plasma tPA and PAI-1 levels [
76]. These results support the notion that interactions among renin–angiotensin, bradykinin, and the fibrinolytic system may play important roles in tPA and PAI-1 biology. Furthermore, it is noteworthy that Ang IV synthesis in the RAAS regulates the release of PAI-1 from endothelial cells when Ang II activation results in an imbalance of ACE1/ACE2 [
77].
PLAT encodes tissue tPA found in endothelial cells that promotes fibrinolysis, exerting its effect by converting plasminogen to plasmin. Plasmin breaks down fibrin polymers, or blood clots, into D-dimer, among other breakdown products. The activity of tPA is inhibited by PAI, which is secreted by endothelial cells, among other cell types and tissues. However, the frequency of the Alu insertion of this gene does not seem to be different between Europeans and Asians. Nevertheless, given the importance of the coagulation/fibrinolysis system in the pathogenesis of COVID-19, the function of the Alu variant of the
PLAT gene cannot be ignored in future studies. tPA is currently being attempted as a therapeutic regimen for various cardiovascular diseases and ARDS associated with COVID-19 [
78].
As described, the relationship between comorbidities and
ACE1 Alu I/D polymorphism has been extensively studied, and there are many reports supporting a positive relationship between their pathophysiology and the DD genotype. It has also been shown that it may be at risk for diseases with higher or lower associations between different genes rather than a single gene [
79]. Dai et al. showed that high levels of ACE1, kallikrein1 (KLK1), and IL-6 were detected in acute myocardial infarction (AMI) patients with the D allele [
80]. Simultaneous increases in Ang II and KLK1 serum levels significantly increased the risk of AMI. Individuals with the
ACE1 DD and
KLK1 GG genotype combination showed a significantly increased risk of AMI compared to individuals with the
ACE1 II and
KLK1 AA genotypes. Although the
KLK1 AA genotype is independent of the Alu, possible interplay of the
ACE1 Alu polymorphism with another gene polymorphism is noteworthy.





{kind=link}