
FISH Map** of Telomeric and Non-Telomeric (AG3T3)3 Reveal the Chromosome Numbers and Chromosome Rearrangements of 41 Woody Plants


Abstract
:1. Introduction
2. Materials and Methods
2.1. Probe and Chromosome Preparation
2.2. FISH Hybridization
2.3. Karyotype Analysis
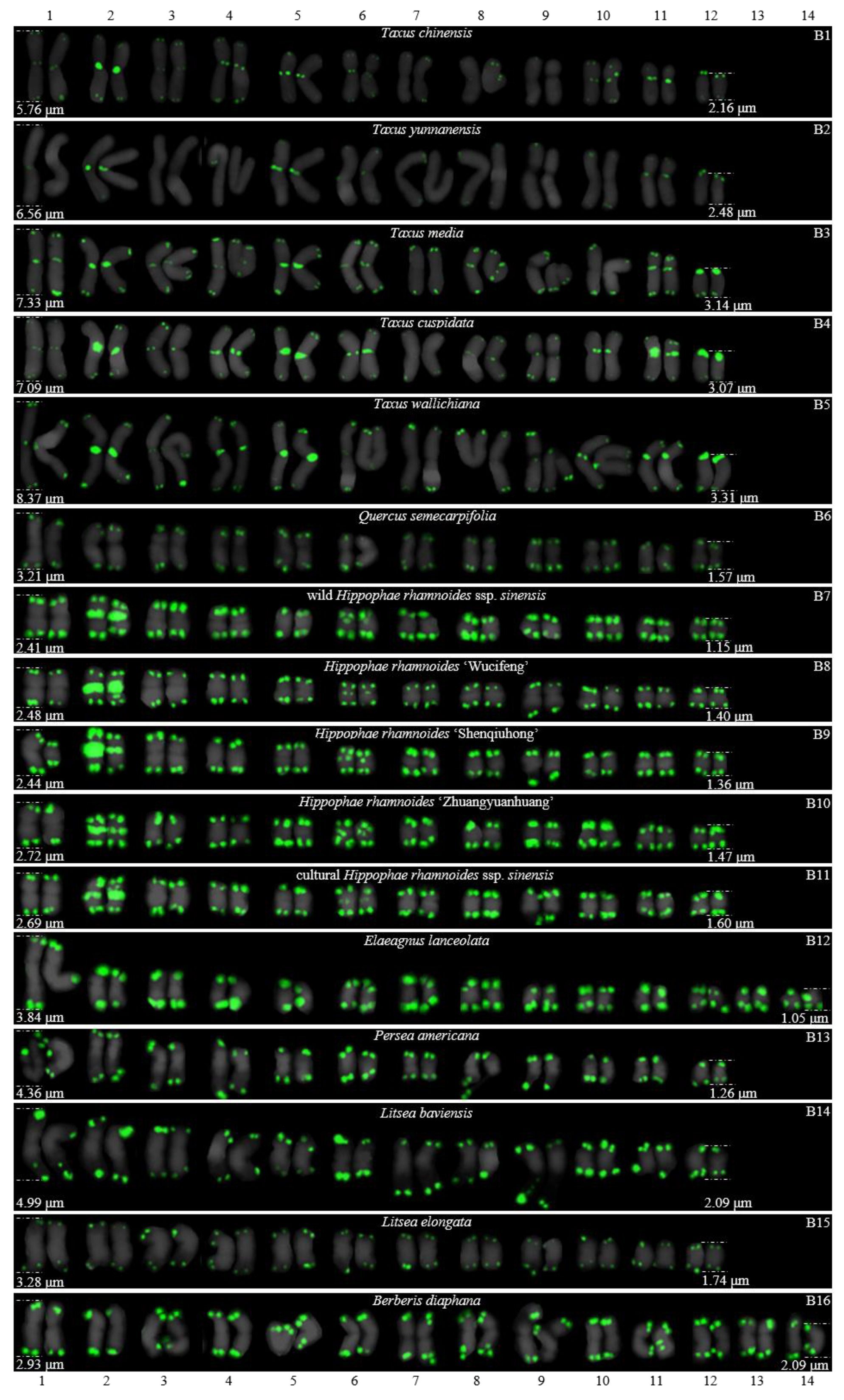
3. Results
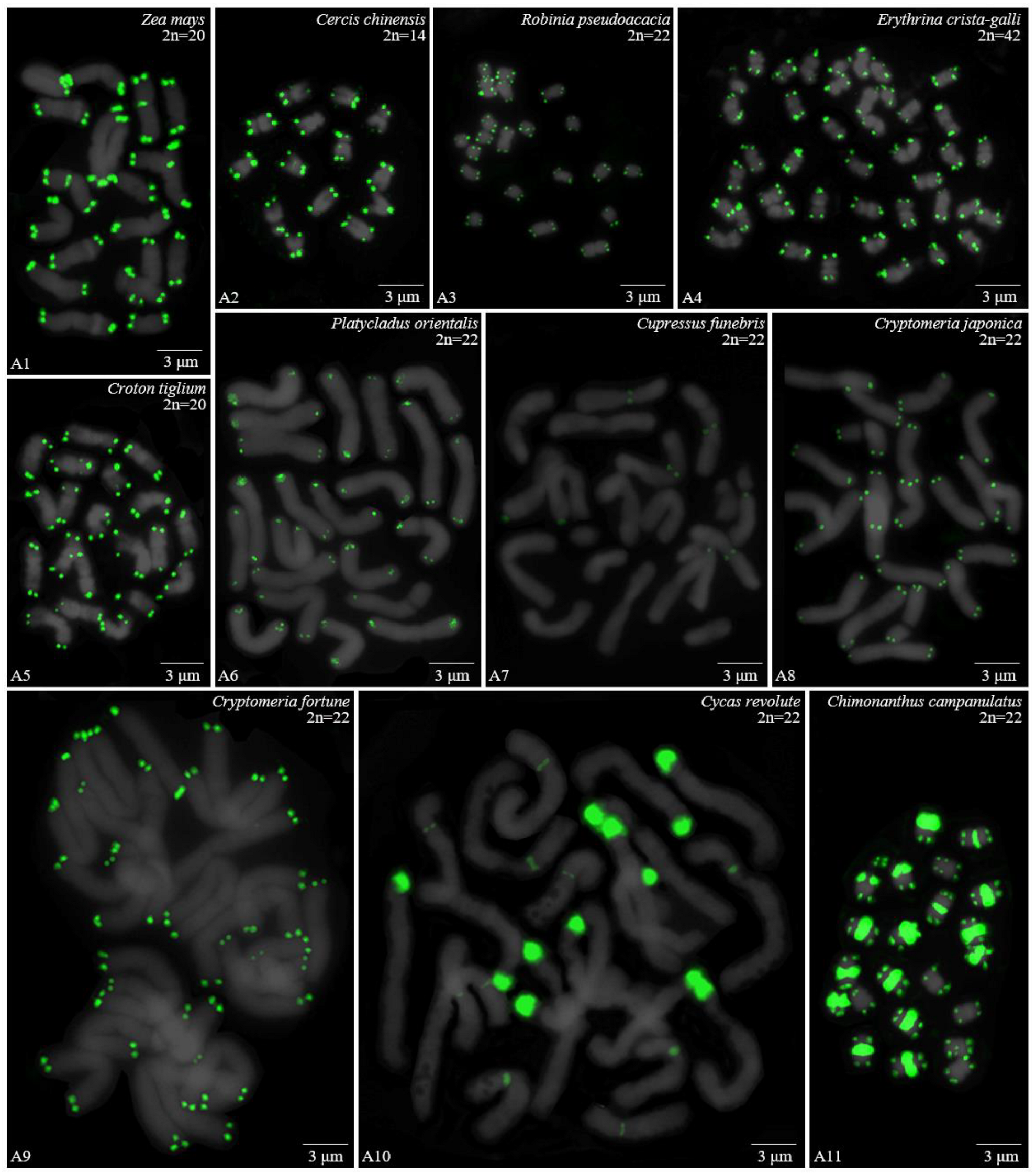
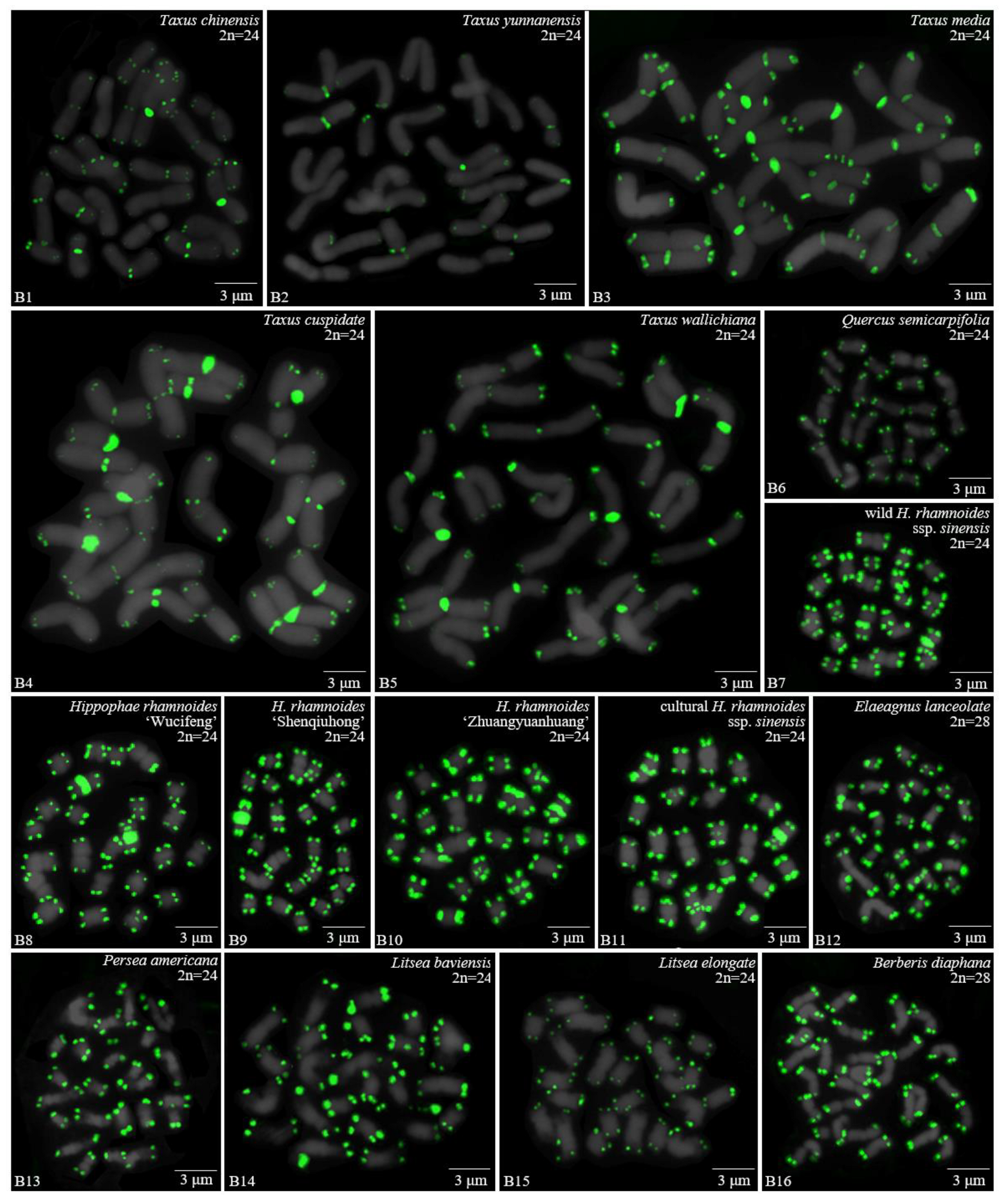
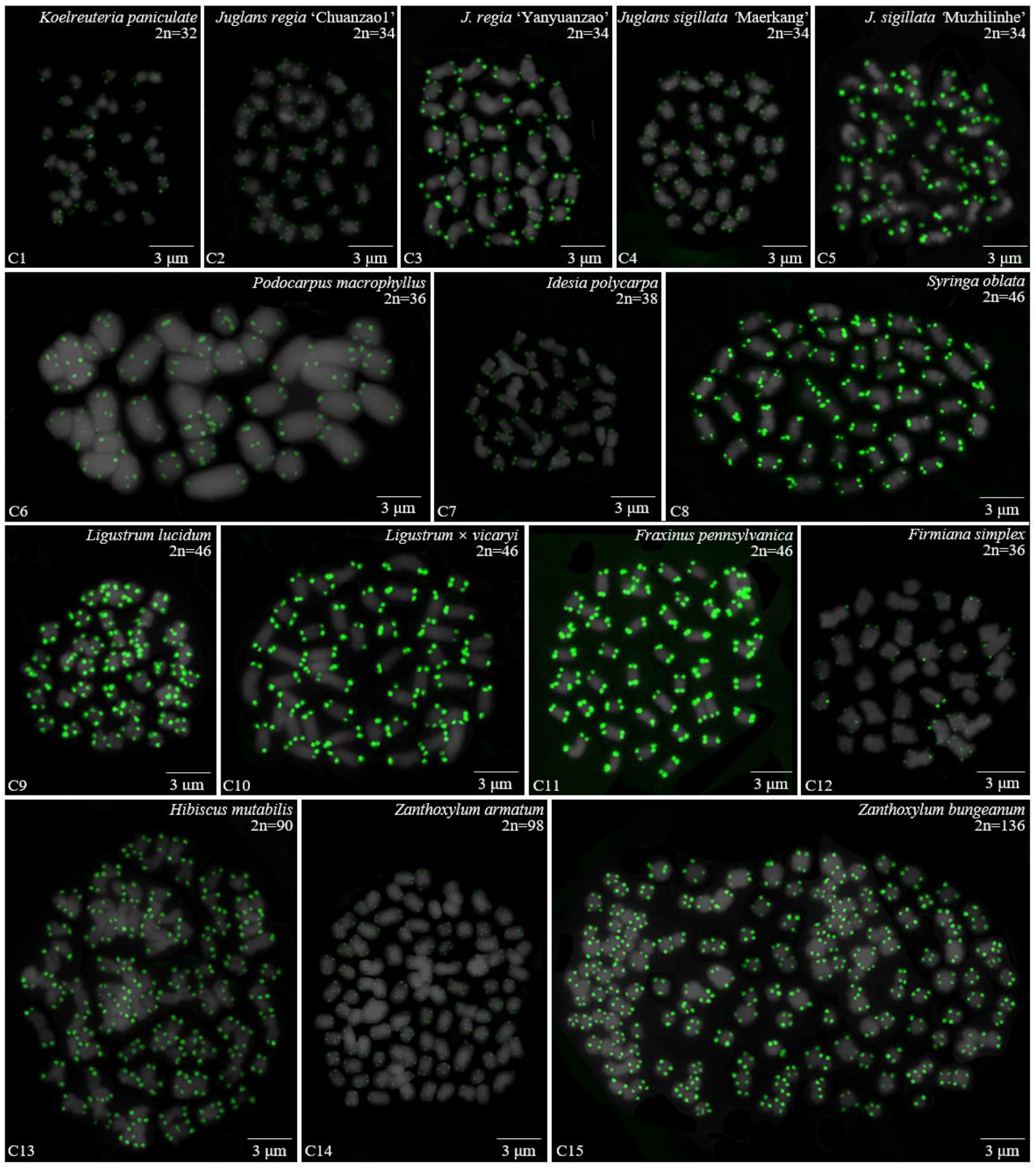
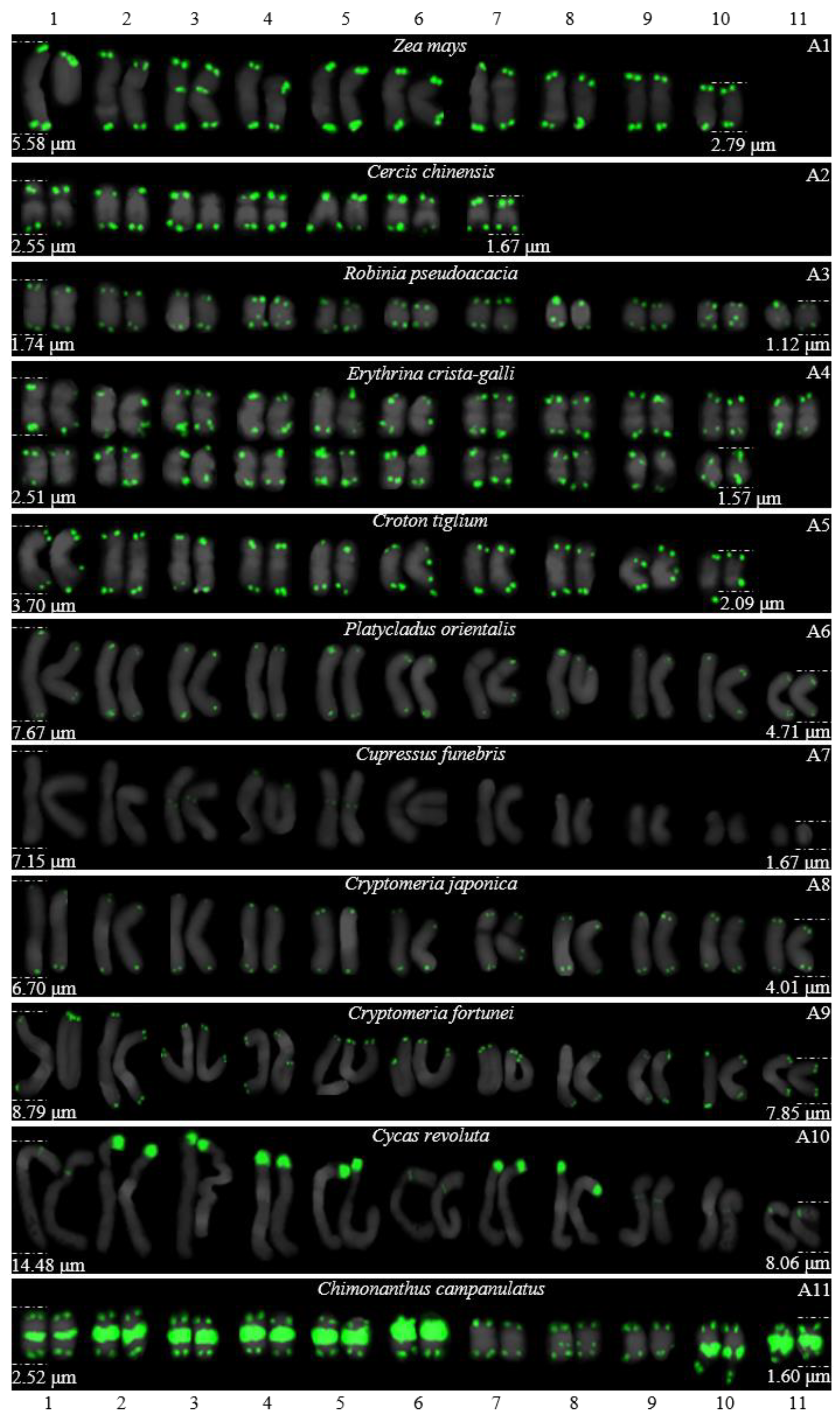
3.1. Karyotype Analysis Revealed Differences among 37 Species
3.2. The Diverse Signal Patterns of (AG3T3)3 Reveal the Complex Genome Architecture
3.3. Proposed Origin of (AG3T3)3 Signal Diversity
4. Discussion
4.1. Karyotype Analysis of Z. mays and 41 Woody Plants
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zakian, V.A. Telomeres: Beginning to understand the end. Science 1995, 270, 1601–1607. [Google Scholar] [CrossRef]
- Wellinger, R.J.; Sen, D. The DNA structures at the ends of eukaryotic chromosomes. Eur. J. Cancer 1997, 33, 735–749. [Google Scholar] [CrossRef]
- Bolzán, A.D. Interstitial telomeric sequences in vertebrate chromosomes: Origin, function, instability and evolution. Mutat. Res. Mutat. Res. 2017, 773, 51–65. [Google Scholar] [CrossRef]
- Traut, W.; Szczepanowski, M.; Vitkova, M.; Opitz, C.; Marec, F.; Zrzavy, J. The telomere repeat motif of basal Metazoa. Chromosome Res. 2007, 15, 371–382. [Google Scholar] [CrossRef]
- Uzlikova, M.; Fulneckova, J.; Weisz, F.; Sykorova, E.; Nohynkova, E.; Tumova, P. Characterization of telomeres and telomerase from the single-celled eukaryote Giardia intestinalis. Mol. Biochem. Parasitol. 2017, 211, 31–38. [Google Scholar] [CrossRef]
- Vitkova, M.; Kral, J.; Traut, W.; Zrzavy, J.; Marec, F. The evolutionary origin of insect telomeric repeats, (TTAGG)n. Chromosome Res. 2005, 13, 145–156. [Google Scholar] [CrossRef]
- Fulneckova, J.; Sevcikova, T.; Fajkus, J.; Lukesova, A.; Lukes, M.; Vlcek, C.; Lang, B.F.; Kim, E.; Eliás, M.; Sykorová, E. A broad phylogenetic survey unveils the diversity and evolution of telomeres in eukaryotes. Genome Biol. Evol. 2013, 5, 468–483. [Google Scholar] [CrossRef]
- Higashiyama, T.; Noutoshi, Y.; Akiba, M.; Yamada, T. Telomere and LINE-like elements at the termini of the Chlorella chromosome I. Nucleic. Acids Symp. Ser. 1995, 34, 71–72. [Google Scholar]
- Derelle, E.; Ferraz, C.; Rombauts, S.; Rouzé, P.; Worden, A.Z.; Robbens, S.; Partensky, F.; Degroeve, S.; Echeynié, S.; Cooke, R.; et al. Genome analysis of the smallest free-living eukaryote Ostreococcus tauri unveils many unique features. Proc. Natl. Acad. Sci. USA 2006, 103, 11647–11652. [Google Scholar] [CrossRef] [Green Version]
- Campomayor, N.B.; Waminal, N.E.; Kang, B.Y.; Nguyen, T.H.; Lee, S.S.; Huh, J.H.; Kim, H.H. Subgenome discrimination in Brassica and Raphanus allopolyploids using microsatellites. Cells 2021, 10, 2358. [Google Scholar] [CrossRef]
- Luo, X.; Liu, J.; He, Z. Oligo-FISH can identify chromosomes and distinguish Hippophaë rhamnoides L. taxa. Genes 2022, 13, 195. [Google Scholar] [CrossRef]
- Petracek, M.E.; Lefebvre, P.A.; Silflow, C.D.; Berman, J. Chlamydomonas telomere sequences are A+T-rich but contain three consecutive G-C base pairs. Proc. Natl. Acad. Sci. USA 1990, 87, 8222–8226. [Google Scholar] [CrossRef] [Green Version]
- Nozaki, H.; Takano, H.; Misumi, O.; Terasawa, K.; Matsuzaki, M.; Maruyama, S.; Nishida, K.; Yagisawa, F.; Yoshida, Y.; Fujiwara, T.; et al. A 100%-complete sequence reveals unusually simple genomic features in the hot-spring red alga Cyanidioschyzon merolae. BMC Biol. 2007, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Sykorova, E.; Lim, K.Y.; Kunicka, Z.; Chase, M.W.; Knapp, S.; Leitch, I.J.; Leitch, A.R.; Fajkus, J. The absence of Arabidopsis-type telomeres in Cestrum and closely related genera Vestia and Sessea (Solanaceae): First evidence from eudicots. Plant J. 2003, 34, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Sykorova, E.; Lim, K.Y.; Fajkus, J.; Leitch, A.R. The signature of the Cestrum genome suggests an evolutionary response to the loss of (TTTAGGG)n telomeres. Chromosoma 2003, 112, 164–172. [Google Scholar] [CrossRef]
- Mravinac, B.; Mestrovic, N.; Cavrak, V.V.; Plohl, M. TCAGG, an alternative telomeric sequence in insects. Chromosoma 2011, 120, 367–376. [Google Scholar] [CrossRef]
- Muller, F.; Wicky, C.; Spicher, A.; Tobler, H. New telomere formation after developmentally regulated chromosomal breakage during the process of chromatin diminution in Ascaris lumbricoides. Cell 1991, 67, 815–822. [Google Scholar] [CrossRef]
- Tran, T.D.; Cao, H.X.; Jovtchev, G.; Neumann, P.; Novák, P.; Fojtová, M.; Vu, G.T.; Macas, J.; Fajkus, J.; Schubert, I.; et al. Centromere and telomere sequence alterations reflect the rapid genome evolution within the carnivorous plant genus Genlisea. Plant J. 2015, 84, 1087–1099. [Google Scholar] [CrossRef] [Green Version]
- Castilho, A.; Vershinin, A.; Heslop-Harrison, J.S. Repetitive DNA and the chromosomes in the genome of oil Palm (Elaeis guineensis). Ann. Bot. 2000, 85, 837–844. [Google Scholar] [CrossRef] [Green Version]
- Červenák, F.; Juríková, K.; Devillers, H.; Kaffe, B.; Khatib, A.; Bonnell, E.; Sopkovičová, M.; Wellinger, R.J.; Nosek, J.; Tzfati, Y.; et al. Identification of telomerase RNAs in species of the Yarrowia clade provides insights into the co-evolution of telomerase, telomeric repeats and telomere-binding proteins. Sci. Rep. 2019, 9, 13365. [Google Scholar] [CrossRef] [Green Version]
- Fajkus, P.; Peška, V.; Sitová, Z.; Fulnečková, J.; Dvořáčková, M.; Gogela, R.; Sýkorová, E.; Hapala, J.; Fajkus, J. Allium telomeres unmasked: The unusual telomeric sequence (CTCGGTTATGGG)n is synthesized by telomerase. Plant J. 2016, 85, 337–347. [Google Scholar] [CrossRef] [Green Version]
- Gomes, N.M.V.; Shay, J.W.; Wright, W.E. Telomere biology in Metazoa. FEBS Lett. 2010, 584, 3741–3751. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, J.; Brandes, A.; Schubert, I. Telomere sequence localization and karyotype evolution in higher plants. Plant Syst. Evol. 1995, 196, 227–241. [Google Scholar] [CrossRef]
- Uchida, W.; Matsunaga, S.; Sugiyama, R.; Kawano, S. Interstitial telomere-like repeats in the Arabidopsis thaliana genome. Genes Genet. Syst. 2002, 77, 63–67. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Liu, J.; Torres, G.A.; Zhang, H.; Jiang, J.; ** of the wild species Phaseolus microcarpus. Genome 2013, 56, 335–343. [Google Scholar] [CrossRef]
- Murat, F.; Xu, J.H.; Tannier, E.; Abrouk, M.; Guilhot, N.; Pont, C.; Messing, J.; Salse, J. Ancestral grass karyotype reconstruction unravels new mechanisms of genome shuffling as a source of plant. Genome Res. 2010, 20, 1545–1557. [Google Scholar] [CrossRef] [Green Version]
- Schweizer, D.; Loidl, J. A model for heterochromatin dispersion and the evolution of C-band patterns. Chromosome Today 1987, 9, 61–74. [Google Scholar] [CrossRef]
- Sousa, G.; Vanzela, A.L.; Crosa, O.; Guerra, M. Interstitial telomeric sites and Robertsonian translocations in species of Ipheion and Nothoscordum (Amaryllidaceae). Genetica 2016, 144, 157–166. [Google Scholar] [CrossRef]
- Islam-Faridi, M.N.; Nelson, C.D.; DiFazio, S.P.; Gunter, L.E.; Tuskan, G.A. Cytogenetic analysis of Populus trichocarpa--ribosomal DNA, telomere repeat sequence, and marker-selected BACs. Cytogenet. Genome Res. 2009, 125, 74–80. [Google Scholar] [CrossRef]
- Lubaretz, O.; Fuchs, J.; Ahne, R.; Meister, A.; Schubert, I. Karyoty** of three Pinaceae species via fluorescent in situ hybridization and computer-aided chromosome analysis. Theor. Appl. Genet. 1996, 92, 411–416. [Google Scholar] [CrossRef]
- Puizina, J.; Sviben, T.; Krajacić-Sokol, I.; Zoldos-Pećnik, V.; Siljak-Yakovlev, S.; Papes, D.; Besendorfer, V. Cytogenetic and molecular characterization of the Abies alba genome and its relationship with other members of the Pinaceae. Plant Biol. 2008, 10, 256–267. [Google Scholar] [CrossRef]
- Rastogi, S.; Ohri, D. Karyotype evolution in cycads. Nucleus 2020, 63, 131–141. [Google Scholar] [CrossRef]
- Shibata, F.; Hizume, M. Survey of Arabidopsis and human-type telomeric repeats using fluorescence in situ hybridization. Cytologia 2011, 76, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Shibata, F.; Matsusaki, Y.; Hizume, M. A comparative analysis of multi-probe fluorescence in situ hybridization (FISH) karyotypes in 26 Pinus species (Pinaceae). Cytologia 2016, 81, 409–421. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.C.; Pellerin, R.C.; Waminal, N.E.; Yang, T.J.; Kim, H.H. Pre-labelled oligo probe-FISH karyotype analyses of four Araliaceae species using rDNA and telomeric repeat. Genes Genom. 2019, 41, 839–847. [Google Scholar] [CrossRef]
- Hizume, M.; Shibata, F.; Matsusaki, Y.; Garajova, Z. Chromosome identification and comparative karyotypic analyses of four Pinus species. Theor. Appl. Genet. 2002, 105, 491–497. [Google Scholar] [CrossRef]
- Islam-Faridi, M.N.; Nelson, C.D.; Kubisiak, T.L. Reference karyotype and cytomolecular map for loblolly pine (Pinus taeda L.). Genome 2007, 50, 241–251. [Google Scholar] [CrossRef]
- Shibata, F.; Matsusaki, Y.; Hizume, M. AT-rich sequences containing Arabidopsis -type telomere sequence and their chromosomal distribution in Pinus densiflora. Theor. Appl. Genet. 2005, 110, 1253–1258. [Google Scholar] [CrossRef]
- Nkongolo, K.K.; Mehes-Smith, M. Karyotype evolution in the Pinaceae: Implication with molecular phylogeny. Genome 2012, 55, 735–753. [Google Scholar] [CrossRef]
- Murray, B.G.; Friesen, N.; Heslop-Harrison, J.S. Molecular cytogenetic analysis of Podocarpus and comparison with other gymnosperm species. Ann. Bot. 2002, 89, 483–489. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; He, Z.J. Distribution of FISH oligo-5S rDNA and oligo-(AGGGTTT)3 in Hibiscus mutabilis L. Genome 2021, 64, 655–664. [Google Scholar] [CrossRef]
- Luo, X.; Chen, J. Distinguishing Sichuan walnut cultivars and examining their relationships with Juglans regia and J. sigillata by FISH, early-fruiting gene analysis, and SSR analysis. Front. Plant Sci. 2020, 11, 27. [Google Scholar] [CrossRef]
- Luo, X.; Liu, J. Fluorescence in situ hybridization (FISH) analysis of the locations of the oligonucleotides 5S rDNA, (AGGGTTT)3, and (TTG)6 in three genera of Oleaceae and their phylogenetic framework. Genes 2019, 10, 375. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Luo, X. First report of bicolour FISH of Berberis diaphana and B. soulieana reveals interspecific differences and co-localization of (AGGGTTT)3 and rDNA 5S in B. diaphana. Hereditas 2019, 156, 13. [Google Scholar] [CrossRef]
- He, Z.; Zhang, W.; Luo, X.; Huan, J. Five Fabaceae karyotype and phylogenetic relationship analysis based on oligo-FISH for 5SrDNA and (AG3T3)3. Genes 2022, 13, 768. [Google Scholar] [CrossRef]
- Luo, X.; Chen, J. Physical map of FISH 5S rDNA and (AG3T3)3 signals displays Chimonanthus campanulatus R.H. Chang & C.S. Ding chromosomes, reproduces its metaphase dynamics and distinguishes its chromosomes. Genes 2019, 10, 904. [Google Scholar] [CrossRef] [Green Version]
- Qi, Z.X.; Zeng, H.; Li, X.L.; Chen, C.B.; Song, W.Q.; Chen, R.Y. The molecular characterization of maize B chromosome specific AFLPs. Cell Res. 2002, 12, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Mu, Y.L.; ** of rDNA and telomeric repeats in 10 Senna species. Hortic. Environ. Biotechnol. 2019, 60, 253–260. [Google Scholar] [CrossRef]
- Waminal, N.E.; Pellerin, R.J.; Kang, S.H.; Kim, H.H. Chromosomal map** of tandem repeats revealed massive chromosomal rearrangements and insights Into Senna tora dysploidy. Front. Plant Sci. 2021, 12, 629898. [Google Scholar] [CrossRef]
- Messier, W.; Li, S.H.; Steward, C.B. The birth of microsatellites. Nature 1996, 381, 483. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | No. | Species | Collection Location |
|---|---|---|---|
| Poaceae | 1 | Z. mays L. | Wenjiang, Sichuan |
| Fabaceae | 2 | Cercis chinensis Bunge | Wenjiang, Sichuan |
| 3 | R. pseudoacacia L. | Wenjiang, Sichuan | |
| 4 | Erythrina crista-galli L. | Wenjiang, Sichuan | |
| Euphorbiaceae | 5 | Croton tiglium L. | Yaan, Sichuan |
| Cupressaceae | 6 | Platycladus orientalis (L.) Franco | Wenjiang, Sichuan |
| 7 | Cupressus funebris Endl. | Wenjiang, Sichuan | |
| 8 | Cryptomeria japonica (L. f.) D. Don | Wenjiang, Sichuan | |
| 9 | Cryptomeria fortune Hooibrenk ex Otto et Dietr. | Wenjiang, Sichuan | |
| Cycadaceae | 10 | Cycas revolute Thunb. | Wenjiang, Sichuan |
| Calycanthaceae | 11 | C. campanulatus R.H.Chang, C.S.Ding | **niu, Sichuan |
| Taxaceae | 12 | Taxus chinensis (Pilger) Rehd. | Wenchuan, Sichuan |
| 13 | Taxus yunnanensis W.C.Cheng and L.K.Fu | Yanan, Sichuan | |
| 14 | Taxus media | Yanan, Sichuan | |
| 15 | Taxus cuspidate Sieb. et Zucc. | Dujiangyan, Sichuan | |
| 16 | Taxus wallichiana Zucc. | Yanan, Sichuan | |
| Fagaceae | 17 | Quercus semecarpifolia Smith | Wenchuan, Sichuan |
| Elaeagnaceae | 18 | wild H. rhamnoides ssp. sinensis Rousi | Wenchuan, Sichuan |
| 19 | H. rhamnoides L. ‘Wucifeng’ | Wenchuan, Sichuan | |
| 20 | H. rhamnoides L. ‘Shenqiuhong’ | Wenchuan, Sichuan | |
| 21 | H. rhamnoides L. ‘Zhuangyuanhuang’ | Wenchuan, Sichuan | |
| 22 | cultural H. rhamnoides ssp. sinensis Rousi | Wenchuan, Sichuan | |
| 23 | Elaeagnus lanceolate Warb. apud Diels | Wenchuan, Sichuan | |
| Lauraceae | 24 | Persea Americana Mill | **chang, Sichuan |
| 25 | Litsea baviensis Lec. | **niu, Sichuan | |
| 26 | Litsea elongate Lec. | **niu, Sichuan | |
| Berberidaceae | 27 | B. diaphana Maxim. | Wenchuan, Sichuan |
| Sapindaceae | 28 | Koelreuteria paniculate Laxm. | Wenjiang, Sichuan |
| Juglandaceae | 29 | J. regia L. ‘Chuanzao1’ | Qingbaijiang, Sichuan |
| 30 | J. regia L. ‘Yanyuanzao’ | Yanyuan, Sichuan | |
| 31 | J. sigillata Dode ‘Maerkang’ | Maerkang, Sichuan | |
| 32 | J. sigillata Dode ‘Muzhilinhe’ | Gulin, Sichuan | |
| Podocarpaceae | 33 | Podocarpus macrophyllus (Thunb.) Sweet | Wenjiang, Sichuan |
| Salicaceae | 34 | Idesia polycarpa Maxim. | Wenjiang, Sichuan |
| Oleaceae | 35 | S. oblata Lindl. | Wenjiang, Sichuan |
| 36 | L. lucidum Ait. | Wenjiang, Sichuan | |
| 37 | L. × vicaryi Rehder | Wenjiang, Sichuan | |
| 38 | F. pennsylvanica Marsh. | Wenjiang, Sichuan | |
| Malvaceae | 39 | Firmiana simplex (Linnaeus) W. Wight | Wenjiang Sichuan |
| 40 | H. mutabilis L. | **niu, Sichuan | |
| Rutaceae | 41 | Zanthoxylum armatum DC. | **yang, Sichuan |
| 42 | Zanthoxylum bungeanum Maxim. | Hanyuan, Sichuan |
| Accession | Species | Chromosome Number | Chromosome Length | Karyotype Asymmetry |
|---|---|---|---|---|
| A1 | Z. mays | 2n = 20 | 2.79–5.58 μm | 2.00 |
| A2 | C. chinensis | 2n = 14 | 1.67–2.55 μm | 1.53 |
| A3 | R. pseudoacacia | 2n = 22 | 1.12–1.74 μm | 1.56 |
| A4 | E. crista-galli | 2n = 42 | 1.57–2.51 μm | 1.60 |
| A5 | C. tiglium | 2n = 20 | 2.09–3.70 μm | 1.77 |
| A6 | P. orientalis | 2n = 22 | 4.71–7.67 μm | 1.63 |
| A7 | C. funebris | 2n = 22 | 1.67–7.15 μm | 4.28 |
| A8 | C. japonica | 2n = 22 | 4.01–6.70 μm | 1.67 |
| A9 | C. fortune | 2n = 22 | 7.85–8.79 μm | 1.12 |
| A10 | C. revolute | 2n = 22 | 8.06–14.48 μm | 1.80 |
| A11 | C. campanulatus | 2n = 22 | 1.60–2.52 μm | 1.58 |
| B1 | T. chinensis | 2n = 24 | 2.16–5.76 μm | 2.68 |
| B2 | T. yunnanensis | 2n = 24 | 2.48–6.56 μm | 2.65 |
| B3 | T. media | 2n = 24 | 3.14–7.33 μm | 2.33 |
| B4 | T. cuspidate | 2n = 24 | 3.07–7.09 μm | 2.31 |
| B5 | T. wallichiana | 2n = 24 | 3.31–8.37 μm | 2.53 |
| B6 | Q. semecarpifolia | 2n = 24 | 1.57–3.21 μm | 2.04 |
| B7 | wild H. rhamnoides ssp. sinensis | 2n = 24 | 1.15–2.41 μm | 2.10 |
| B8 | H. rhamnoides L. ‘Wucifeng’ | 2n = 24 | 1.40–2.48 μm | 1.77 |
| B9 | H. rhamnoides L. ‘Shenqiuhong’ | 2n = 24 | 1.36–2.44 μm | 1.79 |
| B10 | H. rhamnoides L. ‘Zhuangyuanhuang’ | 2n = 24 | 1.47–2.72 μm | 1.84 |
| B11 | cultural H. rhamnoides ssp. sinensis | 2n = 24 | 1.60–2.69 μm | 1.68 |
| B12 | E. lanceolate | 2n = 28 | 1.05–3.84 μm | 3.66 |
| B13 | P. americana | 2n = 24 | 1.26–4.36 μm | 3.46 |
| B14 | L. baviensis | 2n = 24 | 2.09–4.99 μm | 2.39 |
| B15 | L. elongate | 2n = 24 | 1.74–3.28 μm | 1.89 |
| B16 | B. diaphana | 2n = 28 | 2.09–2.93 μm | 1.40 |
| C1 | K. paniculate | 2n = 32 | 0.56–1.12 μm | 2.00 |
| C2 | J. regia L. ‘Chuanzao1’ | 2n = 34 | 0.63–2.60 μm | 4.13 |
| C3 | J. regia L. ‘Yanyuanzao’ | 2n = 34 | 1.01–2.55 μm | 2.52 |
| C4 | J. sigillata Dode ‘Maerkang’ | 2n = 34 | 0.66–1.88 μm | 2.85 |
| C5 | J. sigillata Dode ‘Muzhilinhe’ | 2n = 34 | 0.87–2.79 μm | 3.21 |
| C6 | P. macrophyllus | 2n = 36 | 1.92–7.01 μm | 3.65 |
| C7 | I. polycarpa | 2n = 38 | 0.94–2.23 μm | 2.37 |
| C8 | S. oblata | 2n = 46 | 1.64–2.20 μm | 1.34 |
| C9 | L. lucidum | 2n = 46 | 0.98–1.36 μm | 1.39 |
| C10 | L. × vicaryi | 2n = 46 | 1.37–3.07 μm | 2.24 |
| C11 | F. pennsylvanica | 2n = 46 | 0.98–2.06 μm | 2.10 |
| C12 | F. simplex | 2n = 36 | 1.08–2.37 μm | 2.19 |
| C13 | H. mutabilis | 2n = 90 | 1.22–2.90 μm | 2.38 |
| C14 | Z. armatum | 2n = 98 | 0.98–1.99 μm | 2.03 |
| C15 | Z. bungeanum | 2n = 136 | 0.94–2.62 μm | 2.79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, X.; He, Z.; Liu, J.; Wu, H.; Gong, X. FISH Map** of Telomeric and Non-Telomeric (AG3T3)3 Reveal the Chromosome Numbers and Chromosome Rearrangements of 41 Woody Plants. Genes 2022, 13, 1239. https://doi.org/10.3390/genes13071239
Luo X, He Z, Liu J, Wu H, Gong X. FISH Map** of Telomeric and Non-Telomeric (AG3T3)3 Reveal the Chromosome Numbers and Chromosome Rearrangements of 41 Woody Plants. Genes. 2022; 13(7):1239. https://doi.org/10.3390/genes13071239
Chicago/Turabian StyleLuo, **aomei, Zhoujian He, Juncheng Liu, Hongyi Wu, and **ao Gong. 2022. "FISH Map** of Telomeric and Non-Telomeric (AG3T3)3 Reveal the Chromosome Numbers and Chromosome Rearrangements of 41 Woody Plants" Genes 13, no. 7: 1239. https://doi.org/10.3390/genes13071239