1. Introduction
Oryza sativa L. is a major staple cereal and also serves as a host for several pests that can greatly decrease rice yields [
1,
2]. The small brown planthopper (SBPH),
Laodelphax striatellus (Hemiptera: Delphacidae), is a serious rice sap-sucking insect pest in eastern Asia, including China, Japan and Korea [
3]. Leaves infested with SBPH turn yellow, wilt and even die, reducing yield and quality. Furthermore, SBPH is a potential vector of rice viral diseases, including rice stripe virus and rice black streaked dwarf virus, which also cause significant further yield losses [
4,
5,
6]. Pesticides are commonly used to manage SBPH, which in turn results in the decline of natural enemies, environmental pollution, insecticide resistance and pest resurgence [
7]. As a result, host-plant resistance has been identified as one of the most cost-effective and environmentally safe strategies for managing SBPH [
8,
9]. In recent years, the root microbiome emerged as a vital influence on various plant growth and resistance features [
8,
9]. Beneficial rhizospheric microbes can improve plant health by fixing atmospheric nitrogen, solubilizing plant nutrients which are unavailable in special types of soils such as rock phosphate and enhancing photosynthesis in plants [
10,
11]. Furthermore, these microbes enable the re-generation of damaged plant tissues upon herbivory due to enhanced photosynthetic activities, which in turn stimulates plant tolerance against insect pests [
12,
13,
14]. Greater photosynthesis efficiency enables beneficial microbes to convert more light energy, which allows the generation of induced systemic resistance (ISR) against phloem feeding insects, which can compensate for the losses [
15]. These microbes can also improve plant health in other ways, which include the production of secondary metabolites, enzymes, volatile organic compounds and growth hormones. All these directly or indirectly trigger ISR in plants against insect herbivores [
16]. A rhizospheric bacteria,
Bacillus velezensis YC7010, has antimicrobial, plant growth-promoting and systemic resistance-inducing activity against BPH via rice root drenching [
16]. Colonization of rice roots by
Pseudomonas fluorescens WCS374r induces ISR through enhanced accumulation of phenolic compounds [
17].
P. fluorescens strains Pf1, TDK1 and PY15 display ISR against the leaffolder (
Cnaphalocrocis medinalis) larvae by the activation of PPO in rice plants [
18].
As a biocontrol agent, microbes and their byproducts offer an effective substitute to chemical control, which provide an efficient control with minor or zero ecological impact [
19]. Plant root Rhizobacteria have a latent biological control effect [
20]. This can form part of integrated pest management and has gained attention from researchers as a possible safe and environmentally friendly pest management option [
21]. Recently, microbial insecticides have also attracted considerable attention [
22] because they are more specific, have low relative cost and are more eco-friendly [
23,
24,
25]. The biological control agents derived from actinobacteria,
Streptomyces spp., are reported to be the most researched and well recognized genus of actinomycetes owing to their recognized importance in farming and health, functioning as biofertilizers and biocontrol agents by enhancing plant growth and antiherbivore resistance [
26].
Streptomyces are Gram-positive, culturable, rhizospheric bacteria which produce secondary metabolites and phytohormones [
27,
28,
29,
30,
31,
32].
Streptomyces are well recognized and extensively reported for secondary metabolite synthesis, with a distinct structure and mode of action that provides a key option for biocontrol [
33]. Biological activity of microbial secondary metabolites of actinobacteria, such as
Streptomyces and
Streptoverticillum against
Spodoptera littoralis, caused larval and pupal mortality [
34,
35]. Secondary metabolites from
S. hydrogenans DH16 showed deleterious effects on growth and development of
S. litura larvae. A novel polyketide metabolite isolated from
Streptomyces sp. AP-123 showed larvicidal and growth inhibitory activities against
Helicoverpa armigera and
S. litura [
36]. Similarly, insecticidal activity of crude ethanolic extracts from
Streptomyces sp. have been known to cause mortality of
Sitophilus oryzae larvae [
37].
Angustmycin A was initially extracted from
S. hygroscopicus and an incorrect structure was allocated [
38,
39]. Consequently, the antibiotic decoyinine (DCY) was found to be similar with angustmycin A, and the right structure was determined to be 9-(6-deoxy-β-D-erythro-hex-5-enofuran-2-ulosyl) adenine (
Figure 1) [
40].
Previously, researchers investigated DCY’s effects as a nucleoside antibiotic, antimicrobial and antitumor [
41,
42]. DCY has been referred to as Lingfasu or Wugufengsu in China and its applications in botany and agronomy have been studied extensively [
43]. DCY also promotes the rooting rate, root number and rooting scope of tissue-cultured plantlets of
Fructus momordicae and
Arabidopsis thaliana by increasing the level of endogenous indole-3-acetic acid [
44]. DCY has been shown to enhance rice and maize yields while also improving disease tolerance [
45]. A 50 mg L
−1 DCY solution was applied to the Nanning rice variety “Baixiang 139,” which improved a number of physiological indices such as germination potential, germination rate, root length, bud length, leaf age, seedling base width, plant height, root number, fresh weight and dry matter accumulation [
46]. Though it can affect both auxin and cytokinin regulatory pathways at the same time, it is vulnerable to mutants of auxin and cytokinin targets, suggesting that there could be new approaches to encourage growth and disease resistance [
45]. However, it has only been occasionally recorded that DCY treatment can induce plant insect resistance. The ultimate impacts of DCY on insect performance were determined by the interaction of a positive effect resulting in the form of increased plant growth and a negative effect in the form of induced plant resistance [
47].
Plants normally respond to herbivorous insect feeding with a series of chemical resistance responses. MDA is molecular indicator that shows the injury level of plant cell membrane and higher content of MDA as one of the physiological and biochemical responses of plants against insects [
48]. A variety of stresses (biotic and abiotic) contribute to the rapid synthesis of reactive oxygen species (ROS), which triggers the plant immune system [
49]. Enzymes, such as SOD and CAT, scavenge ROS molecules under steady-state conditions, and they are particularly effective in maintaining ROS balance [
50]. PPO synthesizes insect-resistant secondary metabolites (phenols and polyphenols), and its activity changes in response to insect attack, similar to that of antioxidant enzymes [
48,
51]. The nutritional contents of host plants affect the growth and development of insect herbivores, soluble sugar being one of the most important sources of nutrients for herbivores.
In this study, we tested the hypothesis that DCY treatment could induce a defense response in rice against SBPH. We studied the life cycle of SBPH on DCY treated rice and further investigated whether DCY treatment induces activity changes in rice defense related enzymes (CAT, SOD, POD, PAL and PPO) and MDA, H2O2 and soluble sugars in response to SBPH infestation duration. Such information may provide a better insight into DCY-mediated resistance to SBPH, which could improve rice insect pest management.
4. Discussions
Colonization of host plant roots with beneficial bacteria can induce ISR [
63]. ISR is facilitated via beneficial soil microbes which usually rely on priming [
64]. Priming with microbes is an excellent approach to conserve energy by suppressing basal plant resistance [
65,
66,
67,
68]. Priming can increase resistance against herbivore and pathogen incidence in rice and other crops [
69]. Primed plants demonstrate a rapid and greater cellular defense initiation against insect and pathogen attack in order to enhance plant defense level [
70]. Induced resistance might be utilized as a useful technique in pest management to reduce pesticide applications against insect pests. In particular, induced host plant resistance to insects can be deployed by using chemical elicitors of secondary metabolites that impart insect resistance. Induced response elicitors can be applied to agricultural plants to strengthen the natural defense mechanism against herbivore injury and can serve as a component of integrated pest management for long-term agricultural production [
71].
The present study was initiated with the same concept to evaluate the effects of DCY (secondary metabolites elicitor) on rice against SBPH.
Bacillus velezensis YC7010 is an endophytic bacterium that promotes induced defenses against BPH, when the roots of rice seedlings are treated [
16]. Prior research revealed that a synthetic chemical elicitor (gibberellins, 4-fluorophenoxyacetic acid) boosted rice resistance to piercing–sucking insect pests [
72,
73]. To better understand the impacts of DCY on SBPH, we measured the oviposition performance of SBPH on rice seedlings in the lab and discovered that DCY reduced SBPH fecundity (
Table 1). A previous study also revealed that the exogenous application of abscisic acid promoted rice resistance against BPH by reducing its fecundity [
74].
Azotobacter chroococcum boosts maize yields while decreasing
Mythimna separata pupation rate and fecundity [
75]. The chemical elicitor, gibberellin, had a negative impact on the survival, development and reproduction of BPH female adults [
72]. Although DCY had no effect on SBPH survival, the fecundity of SBPH was significantly reduced in laboratory (
Table 1).
The population life table parameters of SBPH, such as
rm,
R0, DT and λ, were greatly influenced by rice with DCY treatment (
Table 2). Adult emergence, adult longevity and the fecundity of
Spodoptera litura were significantly reduced by
Streptomyces hydrogenans DH16 [
76]. SBPH showed reduced female ratio and fecundity when treated with bacterial pesticide,
Beauveria bassiana sensu lato isolate NJBb2101, which in turn retarded the growth and development of SBPH along with enhanced pesticide susceptibility [
77]. So, we assumed that DCY could enhance rice resistance to SBPH. Insect behaviors and life table parameters were regulated by environmental factors such as humidity, temperature and morphological and chemical components of host plants, including sugars, nitrogen, enzymes and secondary metabolites [
78,
79].
According to our findings, DCY tends to interfere with a variety of defense-related plant defenses that use signaling channels to regulate a variety of physiological and biochemical activities. Excess MDA can damage cell membranes. Conferring to our results, MDA levels in DCY treated plants decreased from 24 to 96 hpi (
Figure 2a). H
2O
2 triggers a series of events in plants that result in the activation of defense genes, defending them against herbivores by activating chloroplast and peroxisome autophagy. H
2O
2 levels in DCY treated rice plants were found to be substantially lower, according to our findings (
Figure 2b). H
2O
2 has been shown to catalyze the accumulation of phenolic polymers in rice parenchyma cells and has been linked to reduced feeding ability in
Sogatella furcifera female adults as well as lower survival rates in other piercing and sucking herbivores [
72].
SOD and CAT can also develop a mechanical barrier to strengthening plant cell wall structures to enhance herbivore resistance [
80,
81]. In this study, DCY boosted the activities of CAT and SOD (
Figure 3a,b), which is parallel with the findings of Harun-Or-Rashid [
16]; he observed that at 48 hpi against BPH, the activities of POD, PAL and PPO were considerably greater in bacteria YC7010 treated rice than in untreated control plants. In our study, the activity of POD was enhanced against SBPH infestation in DCY treated plants with time, and higher significant activity was observed at 96 hpi (
Figure 3c). The same trend of enhanced activities for PAL and PPO was observed in DCY treated rice plants against SBPH (
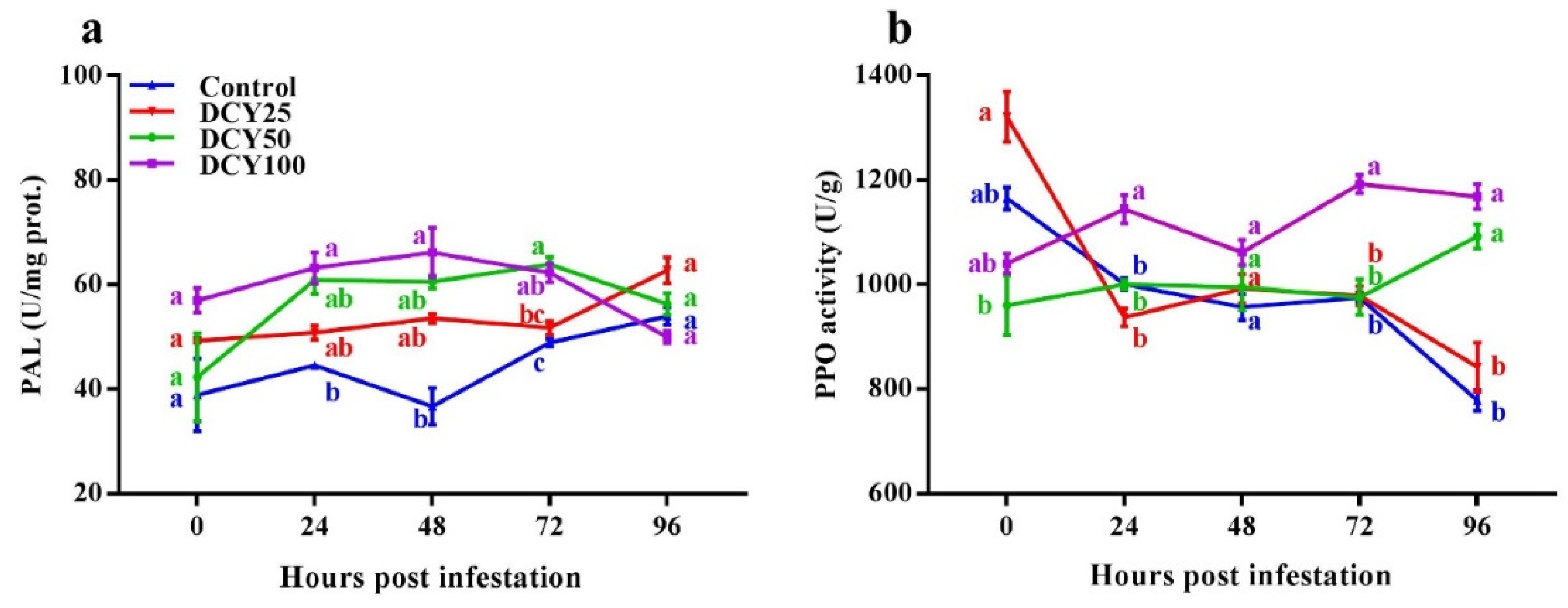
Figure 4a,b). PAL induced resistance against BPH in rice by regulating biosynthesis and accumulation of salicylic acid and lignin via Phenylalanine ammonia-lyase pathway [
82]. In our study, PAL activity was stimulated subsequently on SBPH infestation, and it enhanced significantly in DCY treated plants at 24 hpi (
Figure 4a). PPO catalyzes the oxidation of phenols or polyphenols to synthesize quinones, which can reduce herbivorous insect nutrition absorption. PPO activity was elicited shortly after SBPH infestation, and it increased considerably in DCY treated plants (
Figure 4b).
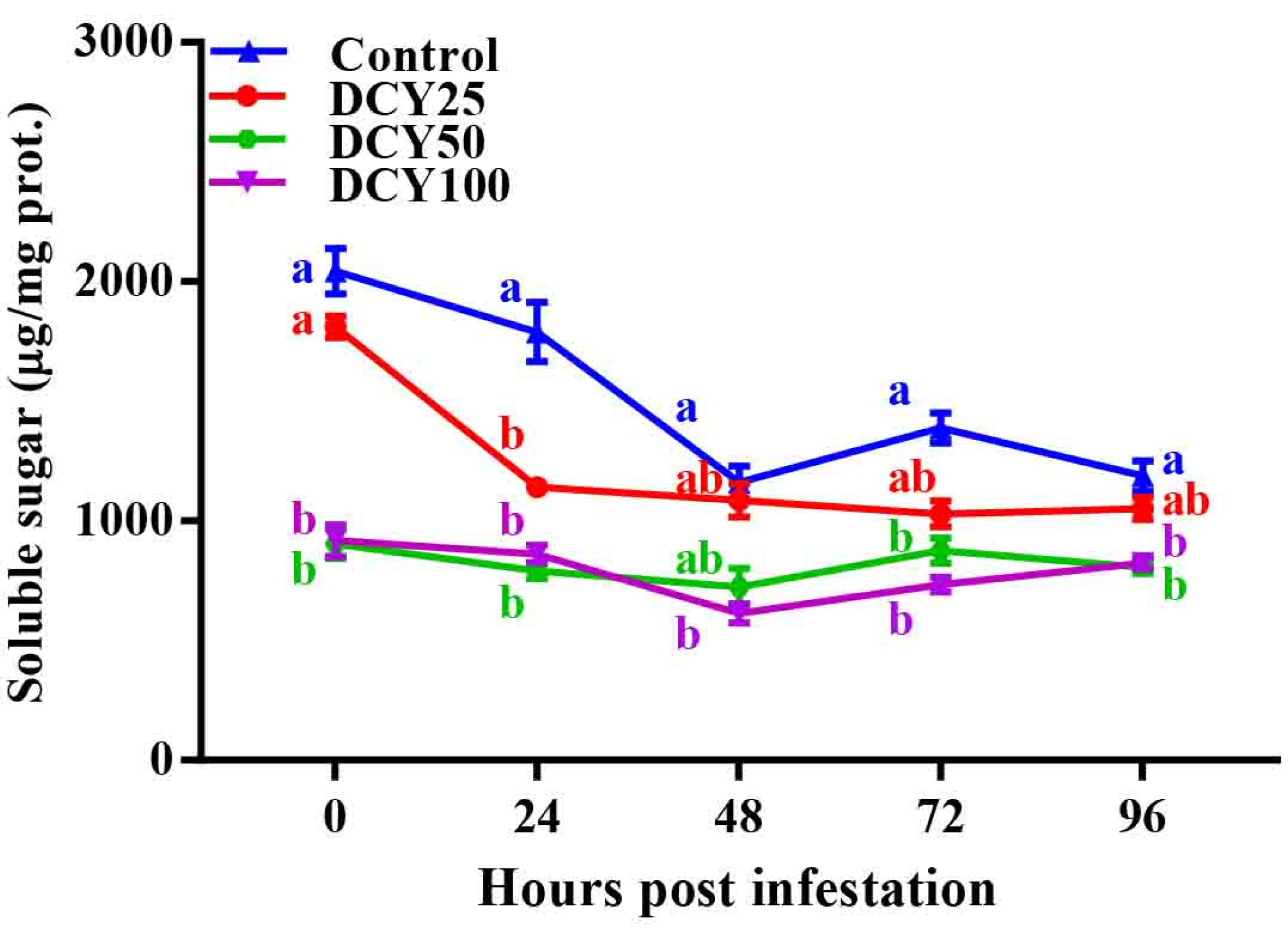
Rice pest resistance is mostly influenced by soluble sugar. In a current study, SBPH infestations have been demonstrated to impact sugar metabolism in rice plants [
83]. The addition of DCY caused a considerable decline in soluble sugar content, according to our findings (
Figure 5). Similar results of low sugar content were detected in the resistant rice variety against higher quantities of total phenols [
84]. In a recent study, the phenolic enzymatic activities were enhanced by DCY treatment. In contrast, **ggangmycin, a fungicide spray, was used to control rice sheath blight (
Rhizoctonia solani), which in turn increased rice plant soluble sugars (sucrose and glucose), sugar metabolism gene expression and resulting BPH population expansion [
85].
In short, the results obtained regarding physiological and biochemical indices of rice plant via GLM showed that all the indices were significantly affected with DCY treatment, while the results regarding SBPH infestation time showed that all the physiological and biochemical indices of rice plant were significantly affected with time, except H
2O
2 (
df = 4, χ
2 = 4.540
, p = 0.338) (
Table 3). A similar approach of GLM analysis was followed to study the mortality of
Rhyzopertha dominica,
Sitophilus oryzae,
Oryzaephilus surinmaneis and
Tribolium castaneum against different phosphine concentrations and insect exposure time intervals [
86]. DCY application could be the possible reason in the priming of ROS, antioxidant enzymes, secondary metabolic enzymes and sugar metabolism mechanisms, which might explain how rice resistance against SBPH is augmented.




 and
and 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}