1. Introduction
Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) has become a globally invasive pest of berries and stone fruits, causing significant economic losses to the fruit industry in its invaded regions [
1,
2,
3,
4]. The native range of
D. suzukii is probably East Asia. It was first described by Matsumura in Japan in 1931 [
1], and has since been reported in eastern China [
5], the Korean Peninsula [
6,
7], Myanmar [
8], Thailand [
9], and other regions in southeastern Asia [
10,
11]. The focus on economic losses due to
D. suzukii in Japan has been concentrated on cherry and blueberry [
1]. However,
D. suzukii has not been considered a serious fruit pest in other southeastern Asia regions, despite recent frequent reports of damage of soft-skinned fruits by
D. suzukii in cherry in China [
12,
13].
In North America,
D. suzukii has invaded most soft-skinned fruit-growing regions in the US and Canada [
2,
14,
15,
16]. The pest caused significant economic losses leading to 100%, 80%, 40%, and 70% losses in cherry, strawberry, blueberry, and raspberry, respectively, in the US in 2008 [
16,
17].
D. suzukii was also detected in Italy, France, and Spain in 2008, and subsequently reported in other European countries [
3,
18]. Furthermore, in 2013, the fly was on the European and Mediterranean Plant Protection Organization (EPPO) A2 list of pests recommended for regulation [
19,
20].
D. suzukii has since been reported in South American and North African countries [
21,
22].
Most
Drosophila species are attracted primarily to damaged, over-ripened, rotted, or fermented fruit; however, the
D. suzukii female adult has a prominent, serrated ovipositor that enables the laying of eggs in ripening or unripe fruits [
23,
24]. The larvae hatch and grow inside the fruit, feeding on the pulp and causing secondary infection by saprophytes [
16], leading to a loss in the quality and commercial value of the fruits.
D. suzukii is a highly polyphagous pest and affects many economically important fruit crops [
25,
26,
27], such as cherry, raspberry, blackberry, blueberry, peach, strawberry, grapes, and other soft-skinned fruits [
28,
29]. In addition, there are more than 60 wild host plants for
D. suzukii, such as
Mahonia aquifolium (Pursh) Nutt. (Berberidaceae),
Frangula purshiana (DC.) A.Gray ex J.G.Cooper (Rhamnaceae), and
Sambucus williamsii Hance (Adoxaceae) [
26].
Insecticides have been widely used in the control of
D. suzukii [
17,
18,
30]. However, spraying insecticides may not kill the larvae feeding inside fruits, and
D. suzukii’s fast development and high reproductive capacity [
31,
32,
33] can result in a rapid population increase [
34,
35]. More importantly, frequent spraying promotes resistance to pesticides [
36], increases insecticide residues on fruits, leads to pest resurgence, affects natural enemies and pollinators [
37], and causes secondary pest outbreaks [
16,
17], affecting control efforts. To safely and effectively control
D. suzukii, the sustainable integrated pest-management (IPM) approach of combining biological, chemical, and cultural control was proposed to reduce the sole reliance on insecticides. Biological control, as a self-perpetuating control option, is an important part of IPM. The introduction of parasitoids to control
D. suzukii population densities provides a new tool for reducing pesticide risk and is an environmentally friendly management strategy in crop fields and non-crop habitats.
China is one of the native ranges of
D. suzukii, and
D. suzukii was first recorded in 1937 [
5]. Parasitoids of
D. suzukii in Northeast China, a major region planting small berries, has not yet been studied. Previous surveys of parasitoids of
D. suzukii have been conducted in Southwestern China (Kunming provinces) [
38], and only limited surveys have been conducted in a few other provinces (Bei**g, Hubei, Sichuan and Jilin) [
38]. To introduce and release natural enemies, especially host-specific and effective parasitoids from the native region of its host pest to the pest’s invaded regions, will be an helpful supplement of IPM. Information on the occurrence and host plants of this pest in its native range is still limited. Therefore, we investigated the occurrence of
D. suzukii and its host plant and associated parasitoids in Northeast China, one of the largest small-berry-planting areas of China. It may provide information on natural enemy recourse for biological control of the pest in Europe, the Americas, and Africa.
4. Discussion
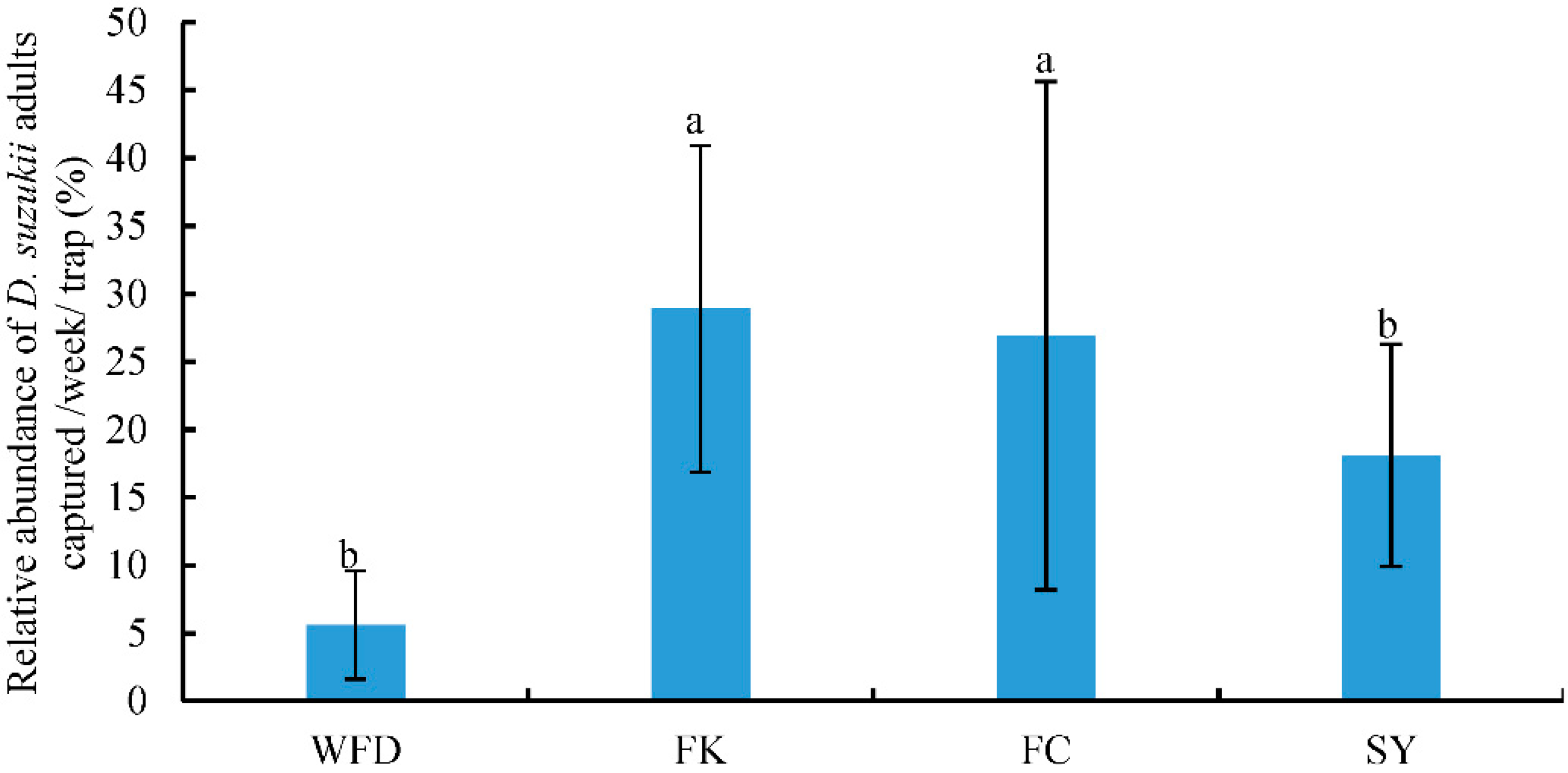
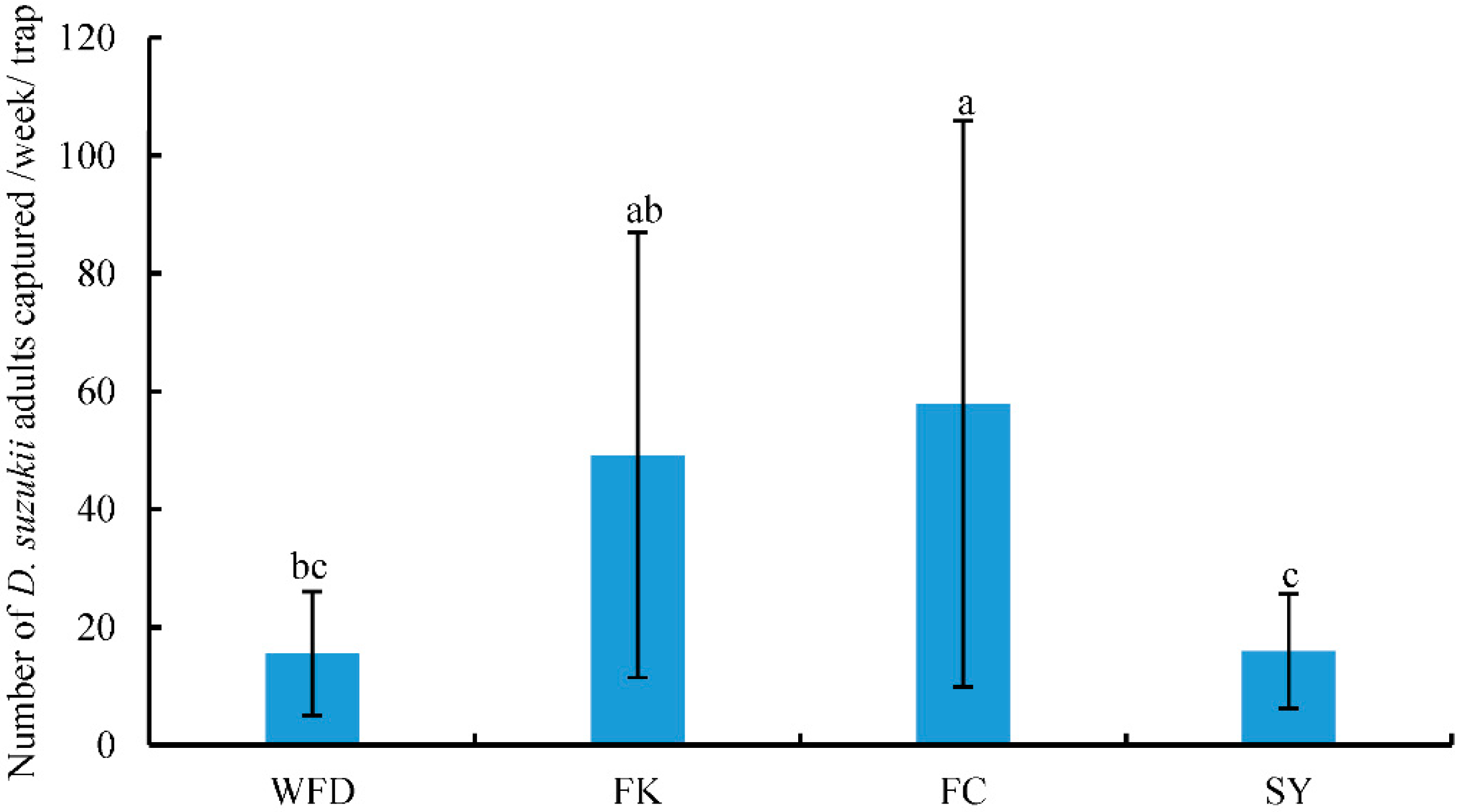
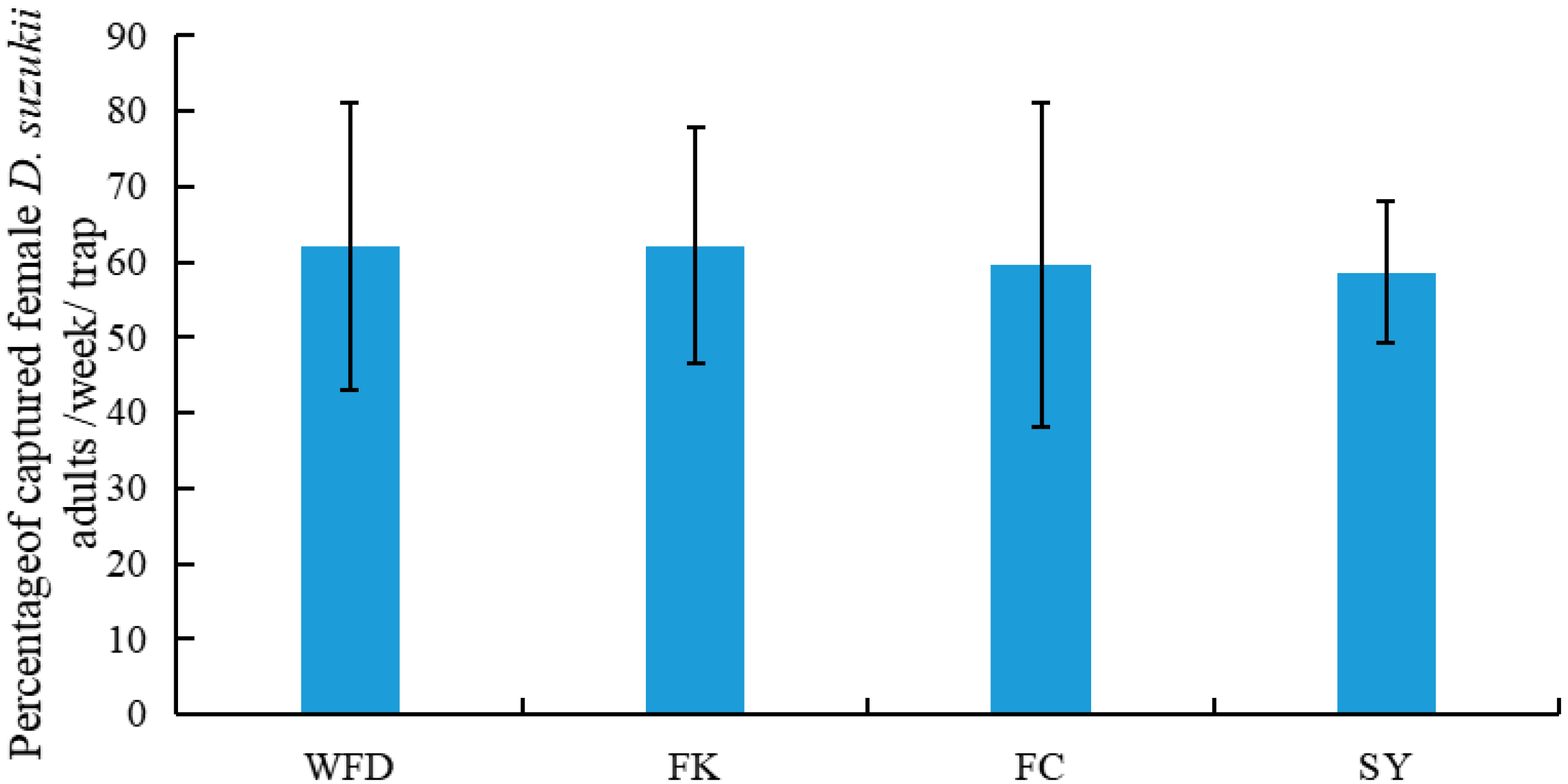
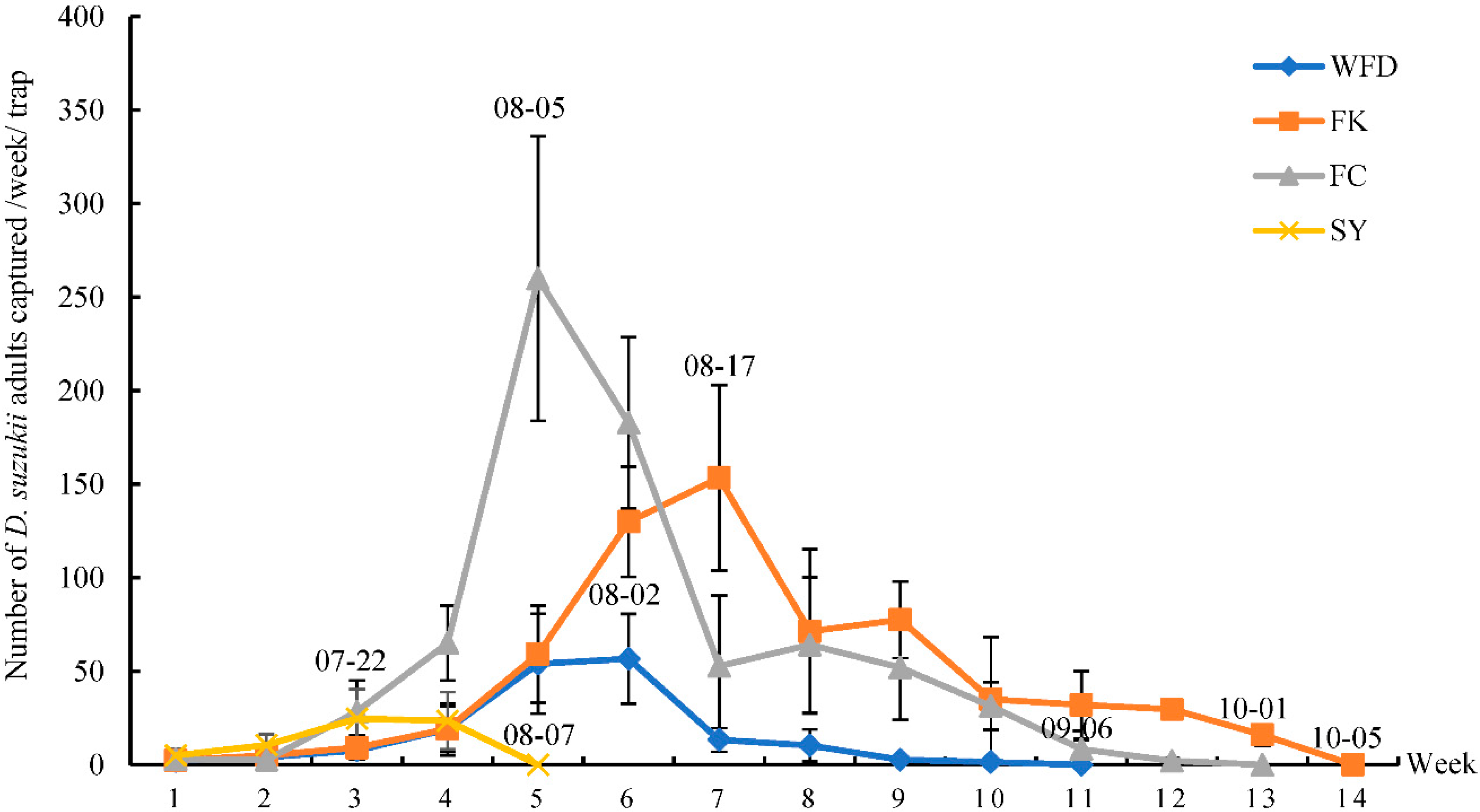
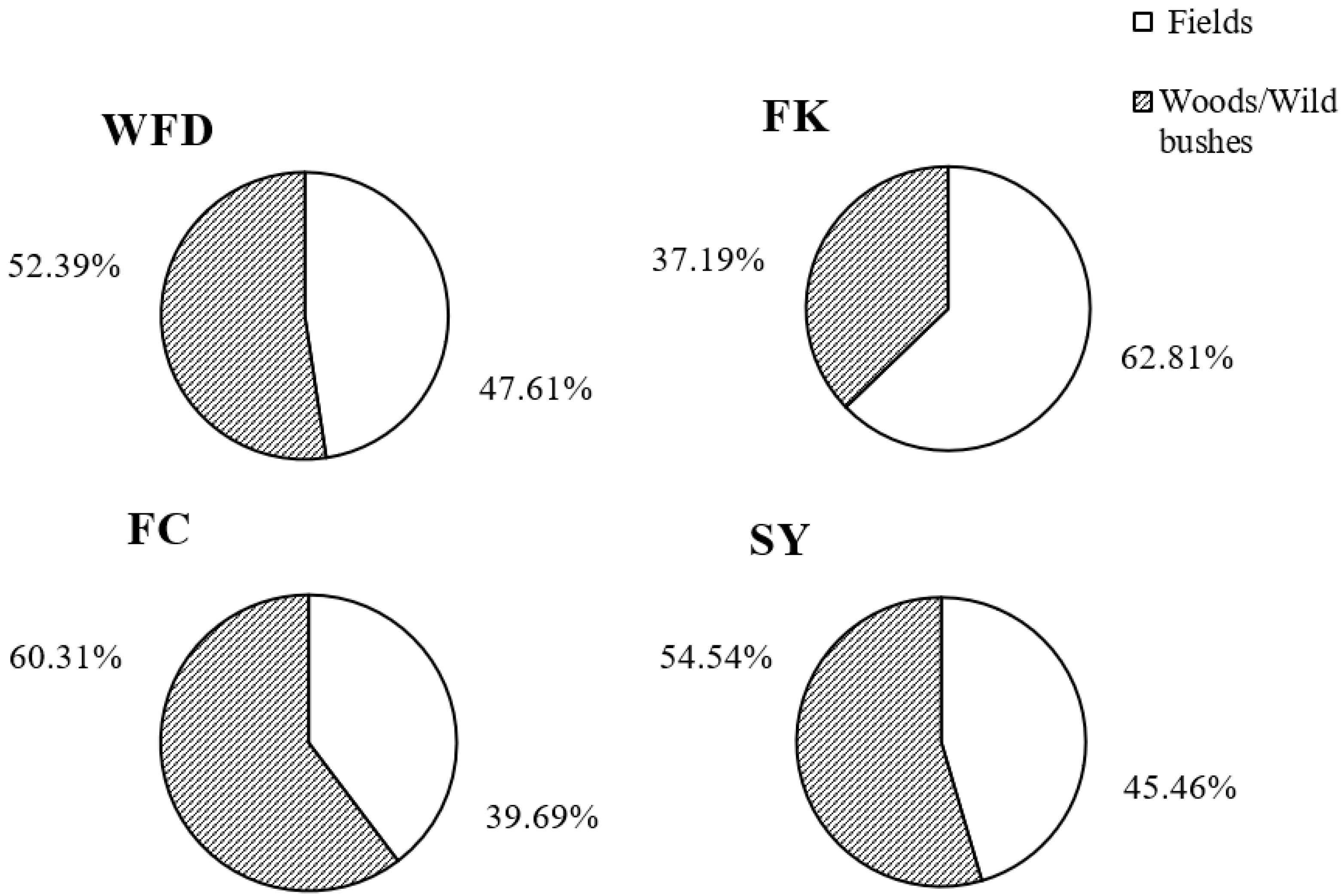
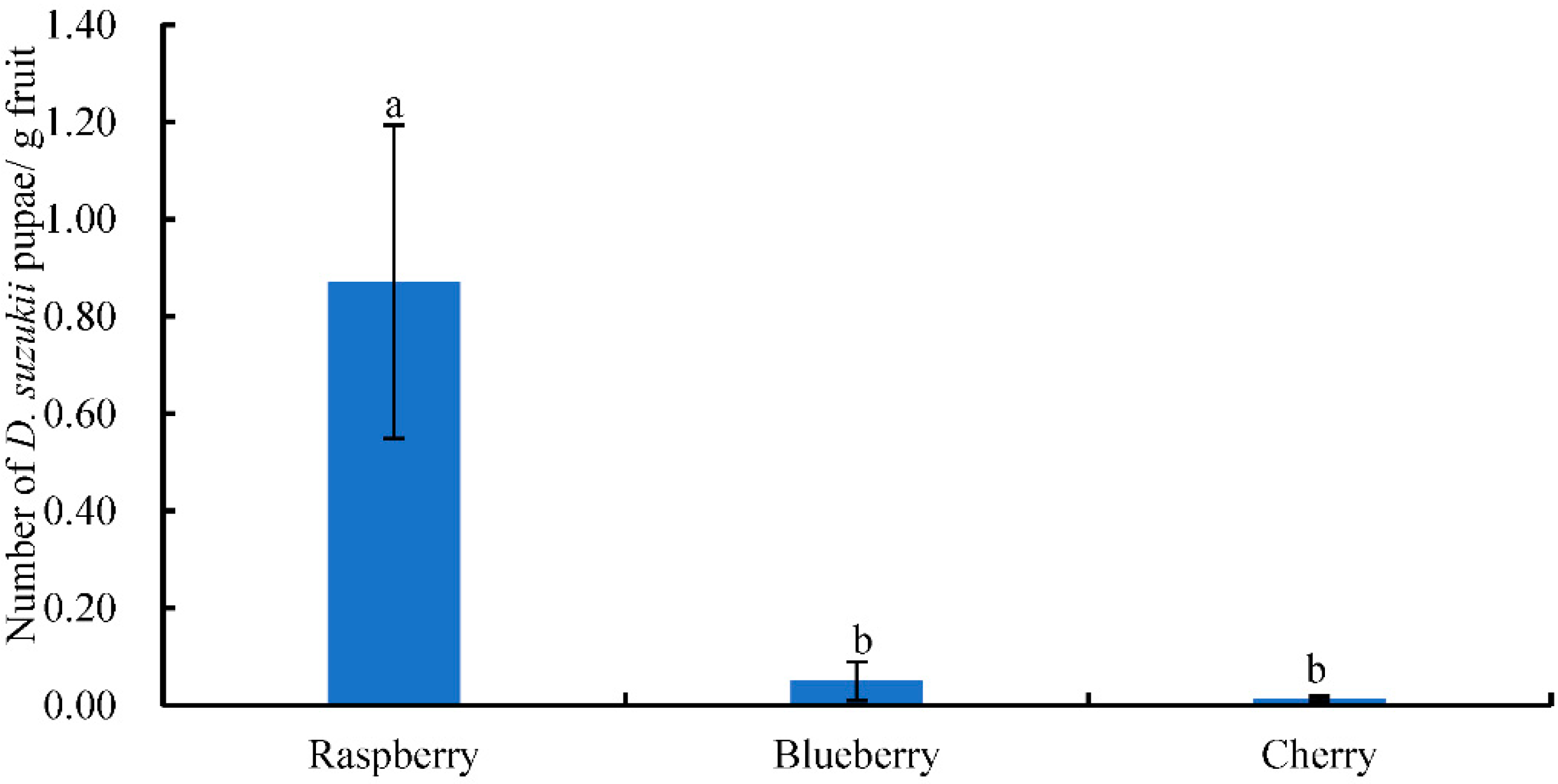
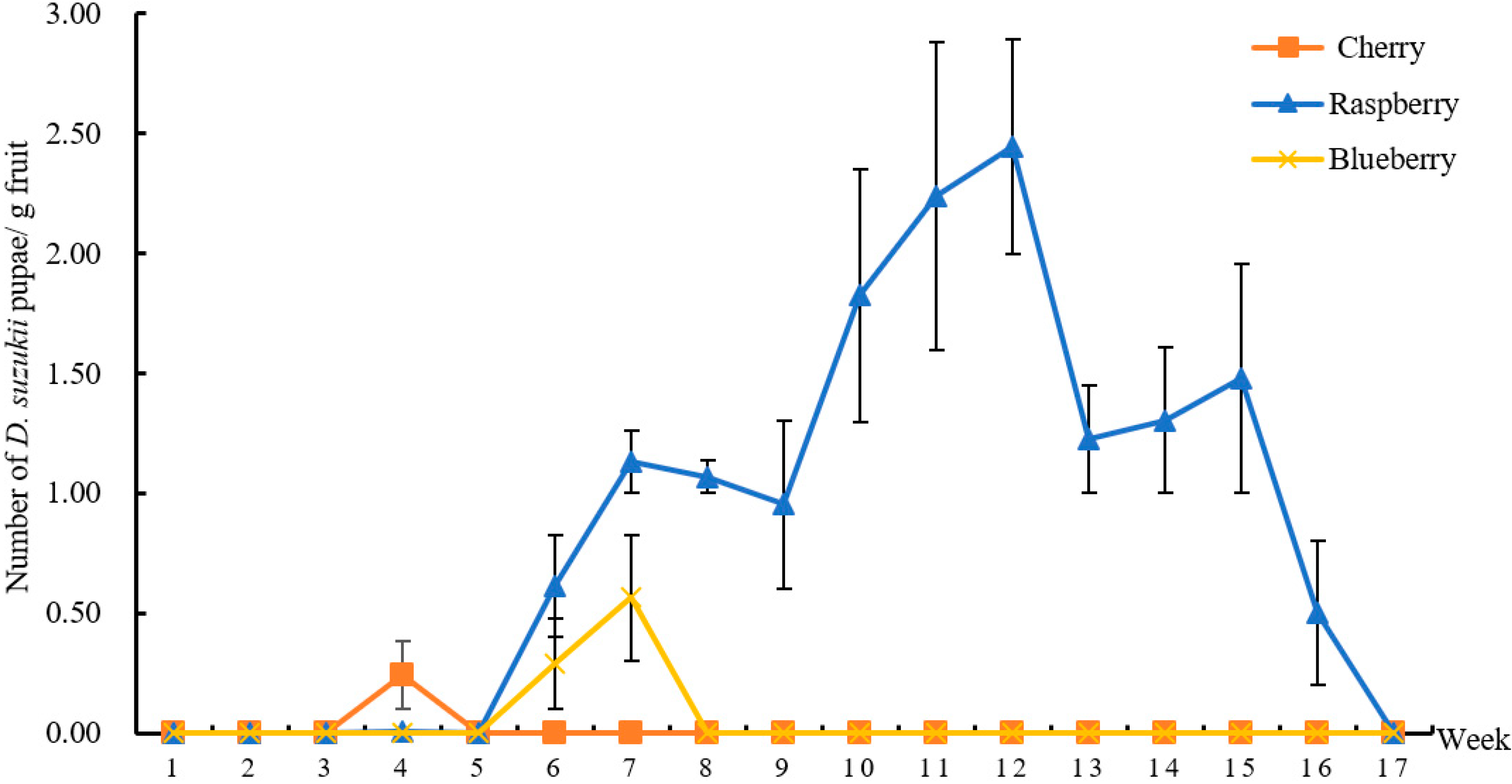
This was the first study that surveyed the occurrence of D. suzukii and its associated host plants and parasitoids in both crop and non-crop habitats in the major berry-fruit-production regions in Northeastern China. Our results showed that: (1) D. suzukii occurred in raspberries, blueberries, and cherries, and raspberries appeared to be the most seriously infested crop; (2) four wild berries from non-crop habitats were also infested by D. suzukii; (3) two species of parasitoids occurred in Liaoning, China; (4) FC had the highest captures of D. suzukii adults in sentinel traps compared to other three survey locations; and (5) throughout the surveyed period (from June to October 2016), the seasonal population dynamics of D. suzukii adults were similar in different survey locations.
Many studies have showed that the number of captured
D. suzukii adults varied among the different geographic locations, the host plant life cycle, temperature, and rainfall [
34,
42,
43]. Like other colder or northern regions in Europe or North America [
44], there was only one peak of adult flies per year in Liaoning. Fly populations appeared in June, increased steadily over the summer to reach a peak in August, and flies were no longer detected by October as temperatures decreased. Among the four locations, the numbers of
D. suzukii adults captured in FK and FC were significantly higher, and collected fruits in these two locations were also more severely damaged than those from WFD and SY. The FK and FC cites were surrounded by woods and bushes, and these non-crop habitats had likely provided source populations of
D. suzukii and affected the occurrence and distribution of the flies. In FK, the number of
D. suzukii larvae collected at the end of the harvest period was higher than that of other periods, probably because those unpicked ripe fruits provided breeding sites for
D. suzukii adults. Furthermore, In FC, the number of
D. suzukii larvae collected in blueberries during the 4th and 5th weeks was significantly higher than in other weeks. This was likely because blueberries were mechanically injured due to the heavy rains and strong winds, and juices from the overripe, damaged, or split fruits could attract
D. suzukii adults.
D. suzukii has a wide range of hosts, and many fruits are oviposition hosts, adult food sources, or provide shelter for overwintering
D. suzukii [
26]. In addition to commonly cultivated soft-skinned fruits,
D. suzukii can also infest a variety of wild or ornamental hosts [
26].
D. suzukii most likely overwinters in forests with wild hosts where refuge and nutrients are more abundant than crop fields. During the spring and early summer, small overwintering populations likely build in non-crop areas to escape monitoring and insecticides [
45,
46]. Surveying and identifying wild hosts, followed by removal of whole plants or fruits, can reduce
D. suzukii habitats and help manage
D. suzukii populations [
44]. The wild berries from ten sampled plant species were collected from non-crop habitats in Liaoning, four of which were infested by
D. suzukii. However, infestations in the field will depend on the level of
D. suzukii populations, host plants (including fruits ripeness, age, and architecture) and relative attractiveness of other hosts in surrounding vicinity [
26]. Therefore, the absence of infestation in the other six fruiting species does not necessarily indicate that they are unsuitable hosts. They are still potential wild hosts for
D. suzukii, and more extensive investigations are needed in the future. Furthermore, studies on host preference by
D. suzukii showed that the fly prefers soft-skinned fruits. Therefore, there is a need to determine the kinds of volatile compounds involved, which will be helpful to develop a better trap than the current yeast traps for the monitoring and control of
D. suzukii in the future.
Many natural enemies, including parasitoids, predators, and entomopathogens, have been evaluated under laboratory conditions for their efficiency against
D. suzukii, and some of them have the potential to be used for biological control of this pest [
47,
48]. In particular, some host-specific parasitoids could be promising natural enemies. There are more than 50 hymenopteran parasitoid species worldwide, attacking
Drosophila species in the larval or pupal developmental stages [
49]. Genetic analyses suggest East Asia is the region of origin for the
D. suzukii populations that invaded North America [
50]. As the origin region of
D. suzukii, East Asia should be the focal region for parasitoid collections [
51]. To date, no locally occurring larval
Drosophila parasitoids can readily develop from
D. suzukii in the invaded regions. In contrast, 19 species of larval
D. suzukii parasitoids were collected from
D. suzukii in East Asia [
48]. Among them,
L. japonica,
Ganaspis brasiliensis (Ihering), and
A. japonica were the dominant parasitoids [
52,
53].
G. brasiliensis and
L. japonica are the most abundant and frequently collected larval parasitoids, and predominantly or exclusively reared from
D. suzukii with reported highest parasitism rates >70% [
54]. At least one genetic group of
G. brasiliensis was found to be the most host-specific to
D. suzukii, which has only been collected from fresh fruits infested by
D. suzukii and other closely related drosophilids [
38,
54]. Thus, the more host-specific
G. brasiliensis was currently being considered for introduction into North America and Europe [
38].
L. japonica was first collected from fresh cherries in Trento, Italy in 2019, and in the coming year,
L. japonica was collected from more locations there, which confirmed that
L. japonica is widely established in the region. They shared more than 99% sequence similarity with specimens of
L. japonica collected in Asia. This means that
L. japonica was probably accidentally introduced into Italy from Asia [
55]. Both
G. brasiliensis and
L. japonica were found established in British Columbia in 2020, probably resulting from accidental introduction [
56]. Although China is one of the native ranges of
L. japonica, there are few studies on it. Previous studies have only confirmed the distribution of
L. japonica in Yunnan, Sichuan, and Bei**g in China [
38]. Further research is needed on the distribution and biological characteristics of
L. japonica in China. As one of the native ranges of
D. suzukii, the Chinese fruit industry has not suffered serious economic losses [
2], probably due to the wide occurrence of some effective native natural enemies of
D. suzukii. At least 10 larval parasitoids,
A. japonica,
Asobara leveri (Nixon) (Hymenoptera: Braconidae),
Asobara mesocauda van Achterberg and Guerrieri (Hymenoptera: Braconidae),
Asobara triangulata van Achterberg & Guerrieri (Hymenoptera: Braconidae),
Asobara pleuralis (Ashmead) (Hymenoptera: Braconidae),
Areotetes striatiferus Li, van Achterberg and Tan (Hymenoptera: Braconidae),
G. brasiliensis,
L. japonica,
Tanycarpa chors Belokobylskij (Hymenoptera: Braconidae), and
Leptopilina sp. (Hymenoptera, Figitidae), and the two pupal parasitoids,
Pachycrepoideus vindemiae (Rondani) (Hymenoptera: Pteromalidae) and
Trichopria drosophilae Perkins (Hymenoptera: Diapriidae) were collected in Sichuan, Yunnan, Bei**g, and Hubei Provinces of China [
38,
53]. The current survey in Liaoning further complemented previous research on the diversity and distribution of these parasitoids in China. However, we only collected
L. japonica and
A. japonica, and did not find
G. brasiliensis. The parasitoid species found in Liaoning were much less diverse than other warmer regions in China as well as in South Korea and Japan [
38,
52,
53]. In the future, more extensive surveys of native parasitoids in East Asia may be needed to discover different species/strains that can establish in different climatic zones in the fly’s invaded regions.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}