
Prevalence, Diversity, and Virulence of Campylobacter Carried by Migratory Birds at Four Major Habitats in China
 ,
,
Abstract
:1. Introduction
2.7. Statistical Analyses
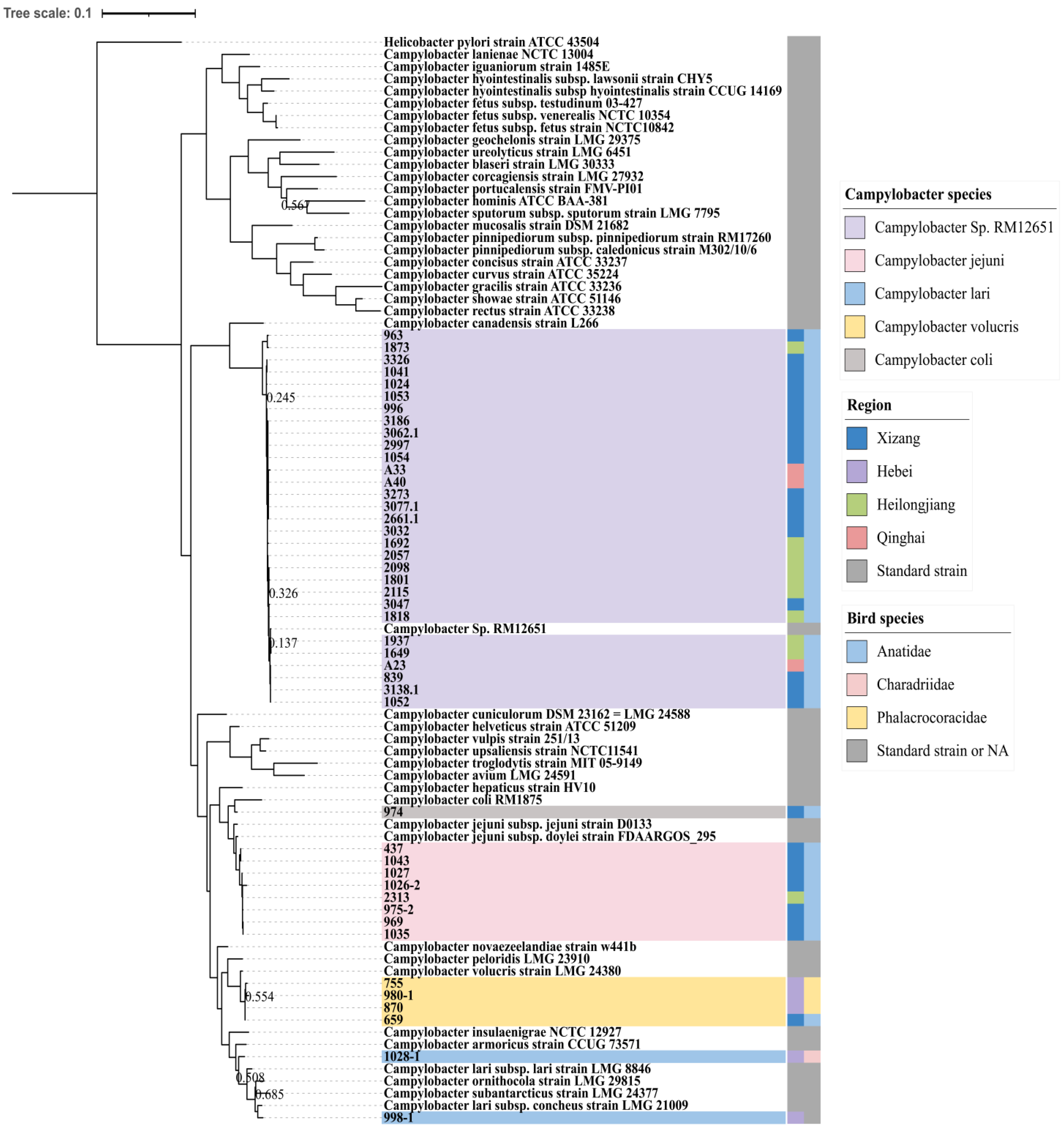
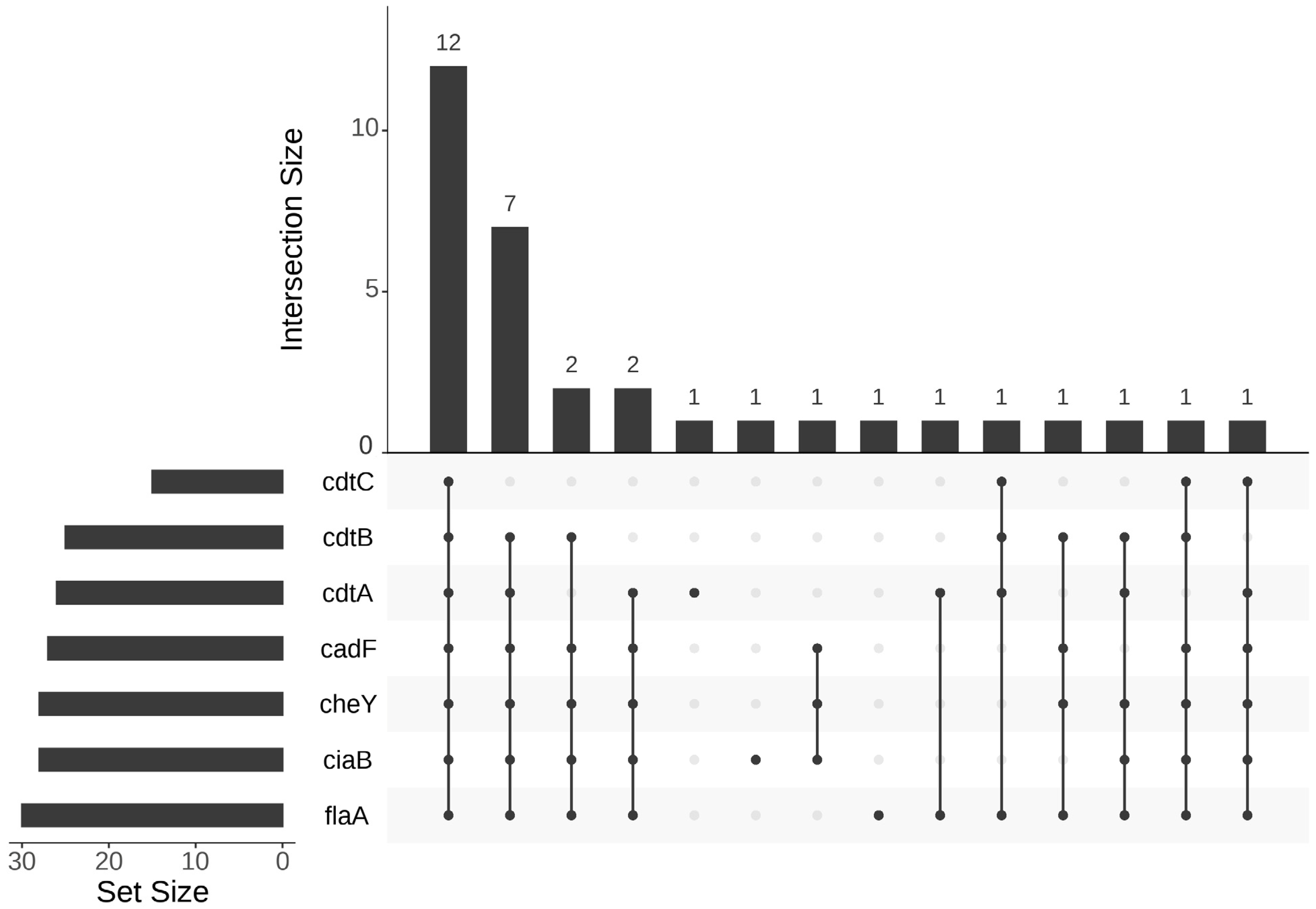
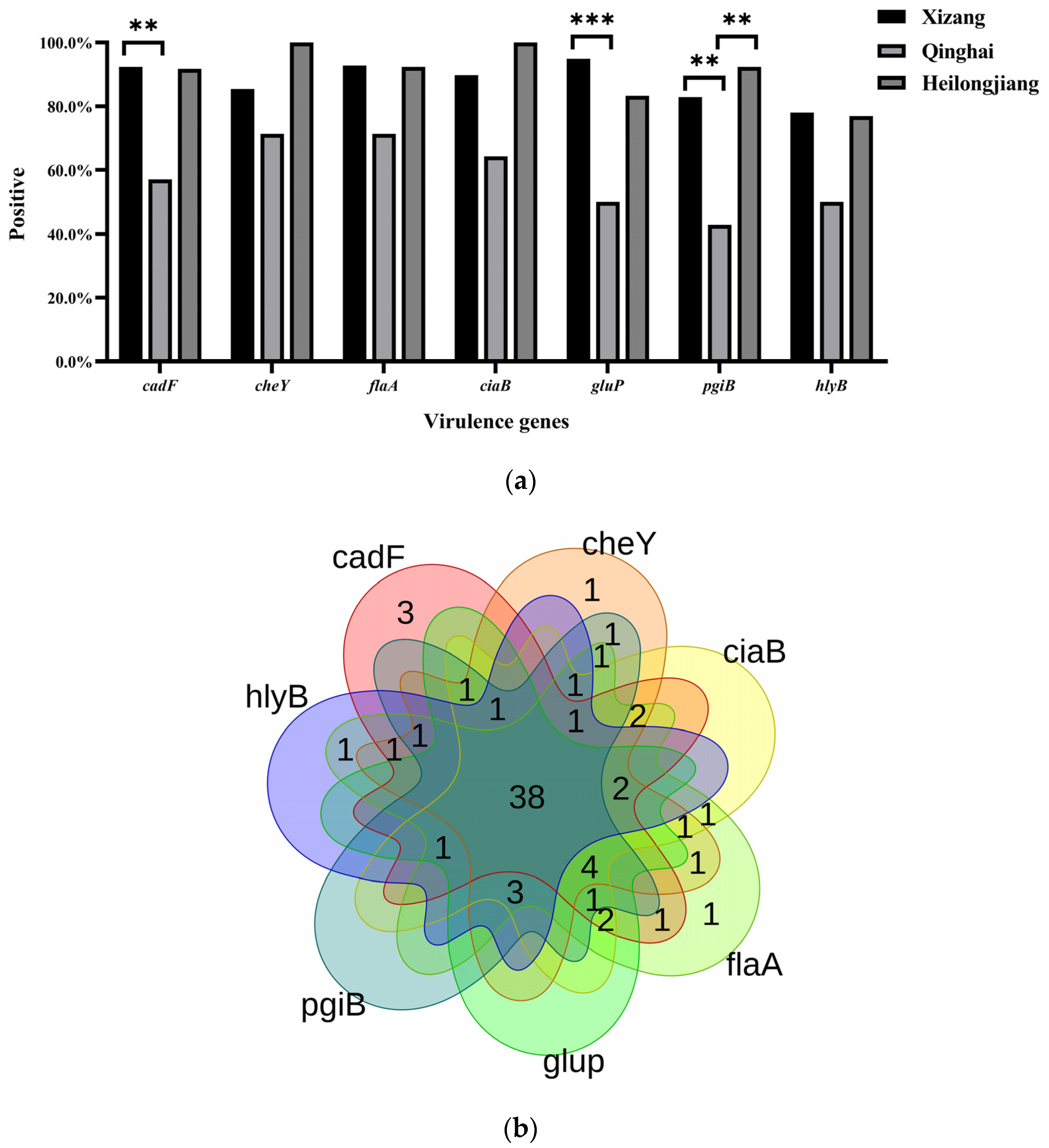
3. Results
3.1. Migratory Bird Types Included in the Current Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campylobacter. Available online: https://www.who.int/zh/news-room/fact-sheets/detail/campylobacter (accessed on 3 February 2024).
- Liu, J.; Platts-Mills, J.A.; Juma, J.; Kabir, F.; Nkeze, J.; Okoi, C.; Operario, D.J.; Uddin, J.; Ahmed, S.; Alonso, P.L.; et al. Use of quantitative molecular diagnostic methods to identify causes of diarrhoea in children: A reanalysis of the GEMS case-control study. Lancet 2016, 388, 1291–1301. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.I.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef]
- Tack, D.M.; Marder, E.P.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Hurd, S.; Scallan, E.; Lathrop, S.; Muse, A.; Ryan, P.; et al. Preliminary Incidence and Trends of Infections with Pathogens Transmitted Commonly Through Food—Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2015–2018. Morb. Mortal. Wkly. Rep. 2019, 68, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Joensen, K.G.; Schjørring, S.; Gantzhorn, M.R.; Vester, C.T.; Nielsen, H.L.; Engberg, J.H.; Holt, H.M.; Ethelberg, S.; Müller, L.; Sandø, G.; et al. Whole genome sequencing data used for surveillance of Campylobacter infections: Detection of a large continuous outbreak, Denmark, 2019. Euro Surveill. 2021, 26, 2001396. [Google Scholar] [CrossRef] [PubMed]
- Lakhan, C.; Badrie, N.; Ramsubhag, A.; Indar, L. Detection of Foodborne Pathogens in Acute Gastroenteritis Patient’s Stool Samples Using the BioFire® FilmArray® Gastrointestinal PCR Panel in the Republic of Trinidad and Tobago, West Indies. Microorganisms 2022, 10, 1601. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Hasegawa, A. Call for Experts on the Microbiological Risk Assessment of Non-Typhoidal Salmonella spp. and Campylobacter spp. in Poultry Meat. Available online: https://www.who.int/news-room/articles-detail/call-for-experts-on-the-microbiological-risk-assessment-of-non-typhoidal-salmonella-spp.-and-campylobacter-spp.-in-poultry-meat (accessed on 14 September 2023).
- WHO. WHO Estimates of the Global Burden of Foodborne Diseases Foodborne Diseases Burden Epidemiology Reference Group 2007–2015. Available online: https://www.who.int/publications/i/item/9789241565165 (accessed on 14 September 2023).
- Silva, W.C.; Targino, B.N.; Gonçalves, A.G.; Silva, M.R.; Hungaro, H.M. Chapter 13-Campylobacter: An Important Food Safety Issue. In Food Safety and Preservation: Modern Biological Approaches to Improving Consumer Health; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 391–430. [Google Scholar]
- Man, S.M. The clinical importance of emerging Campylobacter species. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 669–685. [Google Scholar] [CrossRef]
- Ngobese, B.; Zishiri, O.T.; El Zowalaty, M.E. Molecular detection of virulence genes in Campylobacter species isolated from livestock production systems in South Africa. J. Integr. Agric. 2020, 19, 1656–1670. [Google Scholar] [CrossRef]
- Costa, D.; Iraola, G. Pathogenomics of Emerging Campylobacter Species. Clin. Microbiol. Rev. 2019, 32, 100–128. [Google Scholar] [CrossRef] [PubMed]
- van den Berg, B.; Walgaard, C.; Drenthen, J.; Fokke, C.; Jacobs, B.C.; van Doorn, P.A. Guillain-Barre syndrome: Pathogenesis, diagnosis, treatment and prognosis. Nat. Rev. Neurol. 2014, 10, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Tsiodras, S.; Kelesidis, T.; Kelesidis, I.; Bauchinger, U.; Falagas, M.E. Human infections associated with wild birds. J. Infect. 2008, 56, 83–98. [Google Scholar] [CrossRef]
- Wang, D.; Li, M.; ** system for Campylobacter coli, C. lari, C. upsaliensis, and C. helveticus. J. Clin. Microbiol. 2005, 43, 2315–2329. [Google Scholar] [CrossRef]
- Dingle, K.E.; Colles, F.M.; Wareing, D.R.; Ure, R.; Fox, A.J.; Bolton, F.E.; Bootsma, H.J.; Willems, R.J.; Urwin, R.; Maiden, M.C. Multilocus sequence ty** system for Campylobacter jejuni. J. Clin. Microbiol. 2001, 39, 14–23. [Google Scholar] [CrossRef]
- Poudel, S.; Li, T.; Chen, S.; Zhang, X.; Cheng, W.H.; Sukumaran, A.T.; Kiess, A.S.; Zhang, L. Prevalence, Antimicrobial Resistance, and Molecular Characterization of Campylobacter Isolated from Broilers and Broiler Meat Raised without Antibiotics. Microbiol. Spectr. 2022, 10, e0025122. [Google Scholar] [CrossRef] [PubMed]
- de Melo, A.A.; Nunes, R.; Telles, M.P.d.C. Same information, new applications: Revisiting primers for the avian COI gene and improving DNA barcoding identification. Org. Divers. Evol. 2021, 21, 599–614. [Google Scholar] [CrossRef]
- Rossler, E.; Olivero, C.; Soto, L.P.; Frizzo, L.S.; Zimmermann, J.; Rosmini, M.R.; Sequeira, G.J.; Signorini, M.L.; Zbrun, M.V. Prevalence, genotypic diversity and detection of virulence genes in thermotolerant Campylobacter at different stages of the poultry meat supply chain. Int. J. Food Microbiol. 2020, 326, 108641. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Kang, M.; Jang, H.K. Genetic characterization and epidemiological implications of Campylobacter isolates from wild birds in South Korea. Transbound. Emerg. Dis. 2019, 66, 56–65. [Google Scholar] [CrossRef]
- Miller, W.G.; Yee, E.; Jolley, K.A.; Chapman, M.H. Use of an improved atpA amplification and sequencing method to identify members of the Campylobacteraceae and Helicobacteraceae. Lett. Appl. Microbiol. 2014, 58, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Konkel, M.E.; Talukdar, P.K.; Negretti, N.M.; Klappenbach, C.M. Taking Control: Campylobacter jejuni Binding to Fibronectin Sets the Stage for Cellular Adherence and Invasion. Front. Microbiol. 2020, 11, 564. [Google Scholar] [CrossRef]
- Bolton, D.J. Campylobacter virulence and survival factors. Food Microbiol. 2015, 48, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Young, K.T.; Davis, L.M.; Dirita, V.J. Campylobacter jejuni: Molecular biology and pathogenesis. Nat. Rev. Microbiol. 2007, 5, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.G.; Li, B.Y.; Raghwani, J.; Vrancken, B.; Jia, R.; Hill, S.C.; Fournie, G.; Cheng, Y.C.; Yang, Q.Q.; Wang, Y.X.; et al. Bidirectional Movement of Emerging H5N8 Avian Influenza Viruses Between Europe and Asia via Migratory Birds Since Early 2020. Mol. Biol. Evol. 2023, 40, msad019. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, G.; Li, F.; Ma, T.; Lu, J.; Qian, F. A revised species population estimate for the Bar-headed Goose (Anser indicus). Avian Res. 2017, 8, 7. [Google Scholar] [CrossRef]
- Zhang, J.; **e, Y.; Li, L.; Batbayar, N.; Deng, X.; Damba, I.; Meng, F.; Cao, L.; Fox, A.D. Assessing site-safeguard effectiveness and habitat preferences of Bar-headed Geese (Anser indicus) at their stopover sites within the Qinghai-Tibet Plateau using GPS/GSM telemetry. Avian Res. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Zhou, X.; Li, J.; Yang, H.; Zhang, X.; Wang, M.; Cao, S. Study on the Potential of World Natural Heritage in **ngkai Hu Nature Reserve, Heilongjiang Province. Nat. Herit. 2022, 7, 62–72. [Google Scholar] [CrossRef]
- Hua**, L.; Lu, W. Study on Waterbird Diversity by Season in **ngkai Lake National Nature Reserve, Heilongjiang Province, China. Chin. J. Wildl. 2016, 37, 221–227. [Google Scholar] [CrossRef]
- Fan, Y.; Zhou, L.; Cheng, L.; Song, Y.; Xu, W. Foraging behavior of the Greater White-fronted Goose (Anser albifrons) wintering at Sheng** Lake: Diet shifts and habitat use. Avian Res. 2020, 11, 1–9. [Google Scholar] [CrossRef]
- Jarma, D.; Sánchez, M.I.; Green, A.J.; Peralta-Sánchez, J.M.; Hortas, F.; Sánchez-Melsió, A.; Borrego, C.M. Faecal microbiota and antibiotic resistance genes in migratory waterbirds with contrasting habitat use. Sci. Total Environ. 2021, 783, 146872. [Google Scholar] [CrossRef] [PubMed]
- French, N.P.; Midwinter, A.; Holland, B.; Collins-Emerson, J.; Pattison, R.; Colles, F.; Carter, P. Molecular epidemiology of Campylobacter jejuni isolates from wild-bird fecal material in children’s playgrounds. Appl. Environ. Microbiol. 2009, 75, 779–783. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, K.; Osek, J. Antimicrobial resistance mechanisms among Campylobacter. Biomed. Res. Int. 2013, 2013, 340605. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Sahin, O.; Grover, M.; Zhang, Q. New and alternative strategies for the prevention, control, and treatment of antibiotic-resistant Campylobacter. Transl. Res. 2020, 223, 76–88. [Google Scholar] [CrossRef]
- Liakopoulos, A.; Mevius, D.J.; Olsen, B.; Bonnedahl, J. The colistin resistance mcr-1 gene is going wild. J. Antimicrob. Chemother. 2016, 71, 2335–2336. [Google Scholar] [CrossRef]
- Mohsin, M.; Raza, S.; Roschanski, N.; Schaufler, K.; Guenther, S. First description of plasmid-mediated colistin-resistant extended-spectrum beta-lactamase-producing Escherichia coli in a wild migratory bird from Asia. Int. J. Antimicrob. Agents 2016, 48, 463–464. [Google Scholar] [CrossRef]
- Lin, Y.; Dong, X.; Wu, J.; Rao, D.; Zhang, L.; Faraj, Y.; Yang, K. Metadata Analysis of mcr-1-Bearing Plasmids Inspired by the Sequencing Evidence for Horizontal Transfer of Antibiotic Resistance Genes between Polluted River and Wild Birds. Front. Microbiol. 2020, 11, 352. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Dong, X.; Sun, R.; Wu, J.; Tian, L.; Rao, D.; Zhang, L.; Yang, K. Migratory birds-one major source of environmental antibiotic resistance around Qinghai Lake, China. Sci. Total Environ. 2020, 739, 139758. [Google Scholar] [CrossRef]
- Singh, B.B.; Gajadhar, A.A. Role of India’s wildlife in the emergence and re-emergence of zoonotic pathogens, risk factors and public health implications. Acta Trop. 2014, 138, 67–77. [Google Scholar] [CrossRef]
- Galbraith, P.; Henry, R.; McCarthy, D.T. Plants release, pathogens decease: Plants with documented antimicrobial activity are associated with Campylobacter and faecal indicator attenuation in stormwater biofilters. Sci. Total Environ. 2024, 906, 167474. [Google Scholar] [CrossRef] [PubMed]
- Linton, D.; Owen, R.J.; Stanley, J. Rapid identification by PCR of the genus Campylobacter and of five Campylobacter species enteropathogenic for man and animals. Res. Microbiol. 1996, 147, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.E.; Paccagnella, A.; Law, K.; Melito, P.L.; Woodward, D.L.; Price, L.; Leung, A.H.; Ng, L.K.; Hemmingsen, S.M.; Goh, S.H. Identification of Campylobacter spp. and discrimination from Helicobacter and Arcobacter spp. by direct sequencing of PCR-amplified cpn60 sequences and comparison to cpnDB, a chaperonin reference sequence database. J. Med. Microbiol. 2006, 55, 393–399. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Campylobacter Species | Detection Rate | 16S Identity | Region | |||
|---|---|---|---|---|---|---|
| **zang (n = 82) | Qinghai (n = 14) | Heilongjiang (n = 25) | Hebei (n = 47) | |||
| Campylobacter sp. (RM12651) | 94 (56.0%) | 100.0 (100.0, 100.0) | 51 | 14 | 21 | 8 |
| C. jejuni # | 22 (13.1%) | 100.0 (99.8–100.0) | 19 | - | 1 | 2 |
| C. coli # | 3 (1.8%) | 100.0 (NA) | 3 | - | - | - |
| C. lari | 13 (7.7%) | 100.0 (99.8–100.0) | 3 | - | - | 10 |
| C. volucris | 5 (3.0%) | 100.0 (99.8–100.0) | 1 | - | - | 4 |
| C. novaezeelandiae/armoricus/peloridis/volucris * | 8 (4.8%) | 100.0 (99.7–100.0) | - | - | - | 8 |
| Uncultured bacterium clone * | 17 (10.1%) | 99.6 (97.3–100.0) | 4 | 13 | ||
| Others | 10 (6.0%) | 97.5 (96.6–98.2) | 5 | 3 | 2 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.; Jia, R.; Wang, Y.; Li, J.; Li, Y.; Wang, L.; Wang, Y.; Liu, C.; Jia, E.M.; Wang, Y.; et al. Prevalence, Diversity, and Virulence of Campylobacter Carried by Migratory Birds at Four Major Habitats in China. Pathogens 2024, 13, 230. https://doi.org/10.3390/pathogens13030230
Wu S, Jia R, Wang Y, Li J, Li Y, Wang L, Wang Y, Liu C, Jia EM, Wang Y, et al. Prevalence, Diversity, and Virulence of Campylobacter Carried by Migratory Birds at Four Major Habitats in China. Pathogens. 2024; 13(3):230. https://doi.org/10.3390/pathogens13030230
Chicago/Turabian StyleWu, Shanrui, Ru Jia, Ying Wang, Jie Li, Yisong Li, Lan Wang, Yani Wang, Chao Liu, Elena M. Jia, Yihua Wang, and et al. 2024. "Prevalence, Diversity, and Virulence of Campylobacter Carried by Migratory Birds at Four Major Habitats in China" Pathogens 13, no. 3: 230. https://doi.org/10.3390/pathogens13030230