1. Introduction
On a global scale, salinization of soil is one of the most representative environmental stressors and is a major factor that decreases agricultural productivity and causes a loss of land area of about 2000 ha per day, contributing to 1–2% agricultural soil losses every year worldwide, resulting in decreased crop yield up to 70%, mainly in arid and semi-arid regions, causing a loss of US
$27.3 billion per year [
1,
2,
3]. Concisely, the problem of soil salinization is not only a scourge for agricultural productivity, but also affects soil physicochemical properties and the ecological balance of the area. Several studies have shown that salinity affects almost all factors of plant growth development, including physiological processes, causing nutrient deficiency, reduced vegetative growth, chlorophyll content, inhibited photosynthesis and protein synthesis, and increased susceptibility to phytopathogens [
4,
5,
6,
7]. Antioxidant enzymes are low molecular weight antioxidants produced by plants to confer salt stress tolerance. Remarkably, it has been estimated that the world population will exceed 9 billion people and more than 50% of arable land will be salt-affected due to global climate change by 2050 [
8,
9]. Undeniably, global climate change is a threat in the 21st century and is increasingly becoming a menace to all life on Earth. In contrast, and in response to salinity stress, arable land is declining gradually; hence, it is predicted that agricultural productivity cannot meet the requirements of an exploding world population, which would require an alternative strategy to crop improvement to enhance salt tolerance. Consequently, in recent years, researchers have been trying to investigate the available salinity management techniques to resolve the problem and provide several possible sustainable solutions for salinity reclamation and mitigation.
Endophytic bacteria are ubiquitous in nature, residing inside all parts of plant tissues of most ecosystems and completing their life cycle, mostly in a facultative or obligate symbiotic relationship [
10,
11,
12]. Endophytes are beneficial microorganisms that are important for the development of eco-friendly and sustainable agriculture practices since they help to improve crop yields by enhancing plant growth and protecting crops from microbial diseases [
13,
14,
15]. In recent decades, endophytic actinobacteria have been shown to be attractive natural sources with great potential for the production of bioactive compounds for pharmaceutical and agricultural industries [
16,
17,
18]. Furthermore, beneficial endophytic bacteria have been demonstrated to augment plant growth directly through nutrient solubilization, and the release of plant growth regulators or phytohormones such as indole acetic acid (IAA) enhance a symbiotic nitrogen fixation [
19,
20,
21], as well as reduce infestation by phytopathogens through various mechanisms, including the production of hydrolyzing enzymes such as cellulases and chitinase. The production of siderophores cause degradation of fungal cell walls, aiding in lysis of hyphae and controlling spores germination [
22,
23,
24], as well as the stimulation of systemic acquired resistance and tolerance to abiotic stress factors [
25,
26,
27].
It has been well documented that medicinal plants have been traditionally used worldwide for the treatment of various diseases, and according to the World Health Organization (WHO), about 80% of the world population still relies on medicinal herbs due to their ability to produce a diverse array of biologically active compounds [
28]. The composition of bioactive secondary metabolites synthesized by herbal medicines varies widely depending on the plant species [
24,
29,
30]. Thymus is a traditional medicinal herb in the mint family (
Lamiaceae) and is well known worldwide for its ability to synthesize high quantities of essential oils, which have been widely used in pharmaceutical and food applications [
31,
32,
33,
34]. Individually, beneficial endophytic actinobacteria living within various plant tissues of herbal medicines have the potential for the production of many secondary metabolites with application in agriculture and pharmaceutical industries [
13,
14,
15,
16,
17,
35]. Cotton (
Gossypium) is one of the most important natural fiber crops and is used as an edible oil and biofuel worldwide. However, cotton faces several biotic and abiotic stresses; at present, about three million hectares of cotton in China are infected by the soilborne fungus
Verticillium dahliae, accounting for an annual loss of 10–30% of the cotton yield [
36,
37]. **njiang province, located in the northwest of China, produces about 11% of the global cotton fiber and is disproportionately harmed by Verticillium wilt disease [
38]. Nowadays, the screening of endophytic actinobacteria for their functional role is a promising eco-friendly and bio-economical approach for crop improvements and is gaining prominence [
39,
40]. Thus, the application of salt-tolerant plant growth-promoting (PGP) bacteria to salt- hypersensitive crops, such as maize, bean, tomato, pepper, and cotton, might increase agricultural production during salt-stress conditions [
41,
42,
43,
44,
45]. In a first study of ongoing research, we previously reported the isolation and identification of endophytic actinobacteria associated with wild populations of the Chinese medicinal herb
Thymus roseus Schipcz [
46]. In recent decades, significant advances have been made in the endophytic bacteria interactions with plants. However, studies on the role of endophytic bacteria under stress conditions are still limited. Therefore, the principal goals of the present study were to (1) screen the beneficial endophytic actinobacterial strains associated with wild
T. roseus ex situ; (2) evaluate their ability to stimulate growth-promoting and salt stress tolerance in cotton plants in vivo; (3) evaluate their biological control efficiency against
Verticillium wilt disease in vitro/vivo; and (4) identify the major antimicrobial compounds produced by endophytes in the presence of
V. dahliae, which are likely to be effectors of the antifungal properties. To the best of our knowledge, this is the first report of beneficial plant-microbe interactions associated with the wild medicinal plant
T. roseus to evaluate their potential as biocontrol agents and PGPs in cotton in vitro/vivo.
4. Discussion
Salinity stress is one of the major abiotic stress suppressions limiting crop production in arid and semi-arid regions throughout the world. It has been reported that among the various microbial inoculants, endophytic microorganisms are the most promising beneficial microorganisms for their beneficial exploitation in the field of agriculture [
18,
63]. In particular, endophytic microorganisms can play a significant role as eco-friendly biofertilizers and biocontrol agents for crop disease management [
48,
50,
53]. However, despite the previous work done earlier, there is still limited knowledge on the role of endophytic actinobacteria from different medicinal plants with potential plant growth promotion and antifungal activity [
20,
46,
64]. Therefore, understanding the mechanism underlying plant response to salinity provides new insights into the improvement of salt tolerance crops.
In the present investigation, it has also been documented that endophytic actinobacterial strains exhibited multiple plant beneficial activities, such as the production of IAA (69%), siderophores (84%), phosphate solubilization (79%), and nitrogenase activity (76%). The findings of our study are parallel to some of the previous investigations which reported that endophytes produced several plants promoting traits including the production of phytohormones, siderophores, phosphate solubilization, nitrogenase activity, and cell-wall-degrading enzymes [
48,
65,
66,
67,
68,
69,
70]. In addition, many of the endophytic actinobacterial strains screened in this study were able to produce at least one of the hydrolytic enzymes. The production of cell-wall hydrolyzing enzymes such as cellulose (64%), protease (61%), lipase (62%), and chitinase (34%) are involved in minimizing the challenges imposed by phytopathogens and may play an important role in the biocontrol of plant diseases which have been well-documented previously by various researchers [
26,
48,
53,
71,
72,
73].
Based on the obtained results, the growth of cotton under salt stress was strongly impaired with increasing salinity when cotton was not co-inoculated with selected endophytic actinobacterial strains. In this regard, in response to salinity stress, the maximum decrease recorded in the root, shoot length, and root, shoot weight of cotton were 32.1, 31.5, 29.5, and 33.3%, respectively, at all salt concentrations (
Figure 2). Contrarily, at different salinity levels, the application of selected beneficial plant-microbe
Streptomyces atrovirens XIEG05 showed the strongest stimulation of root length and root weight as compared to the controls. Likewise, the highest increase in the shoot length and shoot weight was obtained by inoculation of
Alloactinosynnema album strain XIEG12, compared to the controls. Accordingly, we observed that the selected salt-tolerant endophytic actinobacterial strains improved the symbiotic performance of cotton under saline conditions. Data obtained in Figure 2 showed that all selected endophytic actinobacteria in this investigation demonstrated a significant role in plant growth activity and displayed increased physiological parameters such as plant biomass and plant length at different salt concentrations. The obtained results are in harmony with many previous studies which suggested that the application of endophytic actinobacteria can increase the growth of different crops such as wheat, rice, lettuce, tomato, pepper, canola, and bean during exposure to salt stress [
74,
75,
76,
77,
78,
79,
80,
81,
82,
83,
84,
85].
Our study showed that the co-cultivation with strain XIEG05 and XIEG12 enhanced cotton growth and salinity tolerance, especially in the development of primary root length, by regulating cell division and differentiation. Apparently, all tested strains appear to influence the root system architecture via root developmental changes, which might enhance plant tolerance to salt stress by the stimulated root system and could explain the enhanced capacity of the plant by solubilizing mineral nutrients and facilitating their availability to plants to acquire and utilize more nutrients which are in agreement with the findings of [
86,
87,
88]. On the other hand, our data recorded that the co-inoculation of XIEG05, XIEG12, and XIEG20 strains induced the accumulation of antioxidant compounds to reduce the impact of salt stress as compared to uninoculated plants (
Figure 3), which is in agreement with the findings of [
24,
48,
89]. For instance, inoculation cotton with strain XIEG05 reduced CAT activity at different salt concentrations compared to uninoculated controls (
Figure 3C); a similar result was observed by [
90,
91]. Moreover, our observations demonstrated that strain XIEG20 decreased the activity of SOD significantly as compared to the uninoculated controls (
Figure 3A). Kadmiri et al. [
92] found in their study that the inoculation of
Pseudomonas fluorescens Ms-01 on wheat grown under salt stress decreased the activity of SOD, which improves the defense pathway. In the present experiment, the highest chlorophyll pigments were observed at all salt concentrations via application of strain XIEG20 as compared to uninoculated plants (
Figure 3D). Our results are in accordance with the findings of Inderbitzin et al. [
93], who reported the effects of water stress and inoculation with beneficial microorganisms on antioxidant status and photosynthetic pigments in basil (
Ocimum basilicum L.).
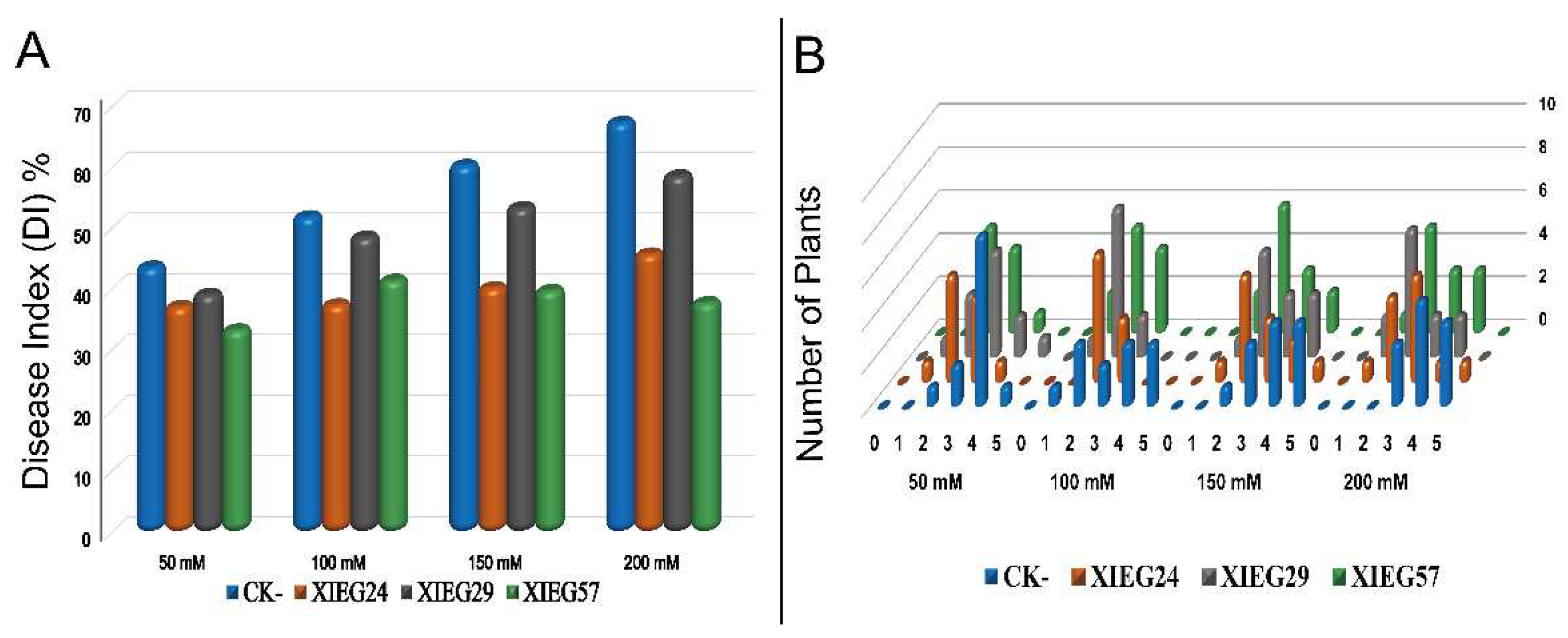
Controlling Verticillium wilt is challenging since the pathogen
V. dahliae can survive in the soil for a long time in dormant forms and causes wilt diseases and crop losses of varying severity, as well as natural ecosystems [
94]. The signs of
Verticillium wilt disease start with yellowing, followed by chlorosis and necrosis of leaves [
53]. In the present investigation, in non-inoculated cotton, salt stress increased the portion of diseased plants up to 43.7, 52.0, 60.4, and 67.5% at 50, 100, 150, and 200 NaCl, respectively, whereas in the presence of strains
Streptomyces albidoflavus (XIEG24),
Curtobacterium flaccumfaciens (XIEG29), and
Nocardiopsis alba (XIEG57) the portion of plants that exhibited disease symptoms decreased at a different level of salt treatments (
Figure 4). These observations demonstrated that the ability of endophytes to colonize internal plant tissues and protect plants from soilborne pathogens was reviewed by [
95]. However, our potential biocontrol strain XIEG57 exhibited the highest disease reduction compared to uninoculated controls. The findings from our study are in agreement with recent reports of [
96,
97,
98], which stated that
Nocardiopsis. sp produces different volatile organic compounds (VOCs) as a biofungicide. Moreover, endophytic actinobacteria associated with medicine plants may play an important role in suppressing the growth of fungal pathogens and can minimize the challenges imposed by phytopathogens including the genus Streptomyces, which are well known for their ability to synthesize different secondary metabolites that play an important role in the biological control of plant disease [
18,
20,
26,
64,
72,
99].
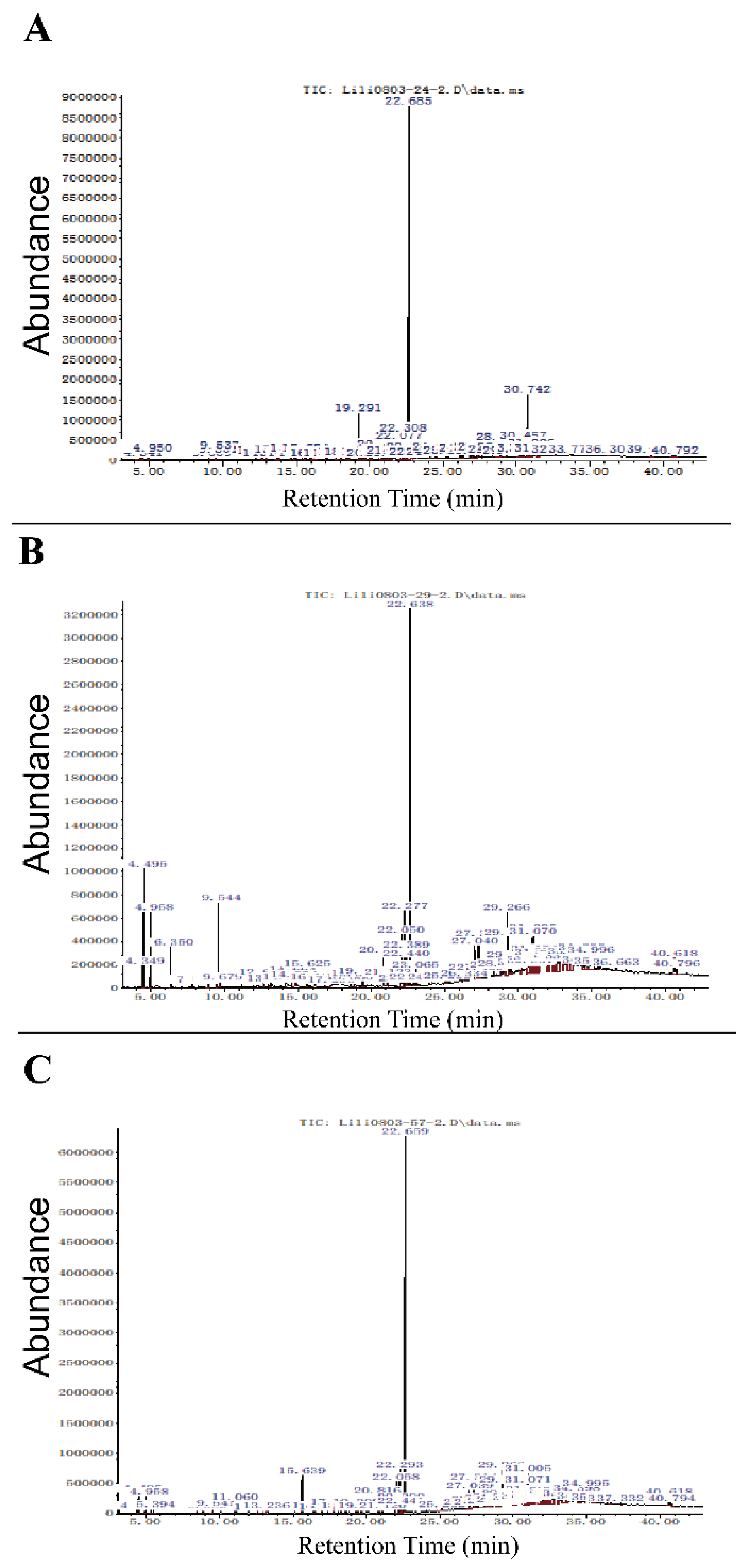
To further understand the underlying mechanism, we conducted Gas-Chromatography/Mass-Spectrometry (GC-MS) analysis to identify the antimicrobial compounds produced by antagonistic strains. Although it is hard to judge whether the components identified by GC-MS are from bacteria or fungi; however, the analysis of obtained data revealed that co-cultures of strains XIEG24, XIEG29, and XIEG57, with
V. dahliae, were tentatively identified as compounds with known antimicrobial, antiphrastic, antitumor, and anticancer properties. Data obtained in (
Figure 5A) showed that the major peaks in cell-free extracts from the co-culture of strain
Streptomyces albidoflavus (XIEG24) are known as antimicrobial compounds such as Dibutyl phthalate [
100], Bicyclo [3.1.1] heptan-2-one [
101].
For strain
Curtobacterium flaccumfaciens (XIEG29), the GC-MS resolved several antimicrobials compounds in the extracts as major peaks such as p-Xylene, Benzene, 1,3-dimethyl-, Benzaldehyde [
102], Phenylethyl Alcohol [
103], and also Phenol, 3,5-dimethoxy-, which is well-known as a polyphenolic compound with antifungal and antibacterial activities [
104]; Diethyl trisulphide [
105], 9-Octadecenamide [
106], Pyrrolo [1,2-a] pyrazine-1,4-dione [
107], 1-Docosene and 1-nonadecene are alkenes [
108], and Nonacos-1-ene [
109] (
Figure 5B).
In this study, about 81 compounds were identified by GC-MS for strain
Nocardiopsis alba (XIEG57); the major peaks in (
Figure 5D) showed antibacterial activity such as Dibutyl phthalate [
100], 1-Docosene, and 1-nonadecene [
108], 1-hexadecene [
110], and antifungal activity against
Pyricularia oryzae such as Pyrrolo [1,2-a] pyrazine-1,4-dione, hexahydro-3-(2-methylpropyl)-, and Nonacos-1-ene [
109], and 13-Docosenamide, (Z)- [
110]. Overall, our results supported the development of natural products that may minimize the need for the application of chemical fertilizer and fungicides, which would be an environmentally friendly approach and preserve biological resources in a sustainable agricultural system.
5. Conclusions
Our study revealed that endophytic actinobacterial associated with medicinal plant T. roseus produced multi-plant growth-promoting substances including IAA, solubilized phosphate, fixed nitrogen, production of siderophores, and produced lytic enzymes (i.e., lipase, cellulase, protease, and chitinase) in vitro. Overall, the greenhouse experiments in this study showed that in response to high salinity stress, inoculation of the selected endophytic actinobacterial strains Streptomyces luteus (XIEG05) and Nocardiopsis dassonvillei (XIEG12) were significantly (p < 0.05) enhanced cotton growth promotion at different salinity levels (50–200) mM compared to un-inoculated cotton. In addition, each tested strain in this study had a different response in terms of cotton growth at certain salt concentrations. In the present study, the activities of antioxidant enzymes (SOD, POD, and CAT) were positively and progressively increased correlated with the salinity level, while the reverse was true for the co-inoculation of cotton with tested endophytic actinobacterial strains as compared with NaCl treatments.
Moreover, the inoculation of cotton with tested endophytic actinobacterial strains resulted in a drastic increase of the chlorophyll synthesis. Noticeably, the results indicated that the three selected antagonistic endophytic actinobacterial strains Streptomyces albidoflavus (XIEG24), Curtobacterium flaccumfaciens (XIEG29), and Nocardiopsis alba (XIEG57) could slow disease development, and the expression of signs was delayed compared to the control plantlets under pathogen-challenged at different salt concentrations. Although all tested endophytic actinobacterial strains conferred some degree of Verticillium wilt resistance, the distribution of disease grades varied dramatically at different salt concentrations compared to uninoculated controls. In addition, most compounds revealed by GC-MS were mainly fatty acid esters, phenols, alkanes, alkenes, and aromatic chemicals and have been reported to have antibacterial and antifungal activity.
In summary, our results provide insights about plant beneficial traits of endophytic bacteria associated with the medicinal plant T. roseus and their interactions with cotton under stressful environments. Moreover, these results support the development of natural products that may minimize the need for the application of chemical fertilizer and fungicides, which would be an environmentally friendly approach and preserve biological resources in a sustainable agricultural system. Further research is needed for future applications in cotton plant growth promotion and crop productivity, as well as to verify the biological control efficacy of selected endophytic bacteria against V. dahliae in the field experiment.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




