1. Introduction
The imbalance of the food rations of the population in megalopolises in the absence of the required amount of essential food micronutrients is a global problem of modern civilization. The deficiency of vitamin A (retinol) was found to be about 10–30%, and that of vitamin E (tocopherol) was 15–35%. This is a major problem in all groups of the population, regardless of age and/or profession. The deterioration of the ecological status of urban areas, the industrialization of the food industry (including the deep processing of the raw materials, long-term storage, refining, and the use of dry substitutes for natural products, etc.) leads to a high necessity for the additional enrichment of the diet with biologically valuable components which are necessary for the prevention of alimentary-dependent diseases [
1].
One of the possible solutions of this problem is to expand the range of functional food products and food additives obtained from the alternative types of plant materials. These plants contain numerous biologically active substances, including natural pigments (carotenoids), α- and β-carotene, fucoxanthin, astaxanthin, lutein, violaxanthin, and zeaxanthin [
2].
The system of conjugated double bonds in carotenoid molecules, as well as structural similarity with some fat-soluble vitamins, determines their high biological value for plants and humans, including their role as a provitamin, their detoxifying activity, the strengthening of the immune system, the chemoprevention of cancer, and the prevention of cardiovascular diseases, cataracts, and other degenerative disorders [
3].
Currently, unconventional raw materials, including microalgae, are considered to be significant sources of carotenoids [
4]. Single-celled green algae
D. salina are known for their ability to accumulate about 0.5 to 10% β-carotene per unit of dry matter [
5]. β-carotene in
D. salina has a high biological activity due to its isomeric composition, which cannot be achieved by chemical synthesis [
6]. The microalgae
Chlorella are considered to be the most valuable, because of their high content of essential amino acids, carbohydrates, vitamins, and lipids, as well as lipid-like compounds, including carotenoids and other valuable plastid pigments [
7,
8,
9,
10]. Algae contain xanthophylls of a unique chemical structure. This is characteristic of the individual taxa, as algae’s oxycarotenoids are derivatives of not only β- but also α- and γ-carotenes. Thus, algae of the
Chlorophyta group contain special pigments: phycocyanin, siphonoxanthin, astaxanthin, loroxanthin, zeinoxanthin, and neoxanthin, as well as allene and acetylenic carotenoids such as diatoxanthin and diadinoxanthin, and many others.
Depending on the cultivation conditions, the nutrient composition of the Chlorella cell can vary significantly. Taking into account the specificity of the Chlorella pigments’ application and the variety of directed cultivation techniques, it is possible to stimulate the biosynthesis of secondary carotenoids in order to isolate and apply them further for nutraceuticals and para-pharmaceuticals.
The effect of the culture medium’s composition and bubbling on the quantitative accumulation of pigments, including carotenoids, in the microalgae
Chlorella vulgaris was studied [
11]. Due to their high adaptive activity, unicellular algae are able to synthesize a large number of secondary carotenoids under various stress conditions, e.g., the osmotic shock of nitrogen and/or phosphorus deficiency, a high intensity of absorbed light, non-optimal temperatures, and the presence of reactive oxygen species in the culture medium or their synthesis. The connection between the presence of reactive oxygen species in the culture medium and the biosynthesis of astaxanthin by heamatococci was proven [
12]. Therefore, it is possible to stimulate the biosynthesis of secondary carotenoids with a simple variation in the cultivation conditions.
An equally important element of the carotenoid production technology is the preliminary activation of the obtained biomass. Despite the fact that the cell membrane of the microalgae
C. vulgaris of the planktonic strain has a thinner membrane than the other species of
C. vulgaris, the homogenization of the biomass has a positive effect on the yield of the final product [
13].
High significance is attached to the method of carotenoid extraction from the microalgae biomass. Different solvents are used to extract carotenoids from the biomass. For example, ethanol and 2-methyltetrahydrofuran (MTHF) were used to maximize the extraction of individual carotenoids [
14] at a temperature of 110 °C, where 45% of all of the carotenoids were extracted. A mixture of MTHF and ethanol in a 1:1 ratio from the raw biomass led to an increase of the extractable carotenoids to 66%. However, MTHF is quite toxic, and cannot be used to produce a dietary supplement.
Microalgae represent a successful model for the commercial production of carotenoids. In addition, the global carotenoid market is projected to reach US
$ 1.7 billion by 2022, with the largest shares being in lutein, astaxanthin, and β-carotene [
15].
The aim of this work is to investigate the modes of directed cultivation of Chlorella microalgae in order to obtain biomass with a high content of carotenoids, and to develop a method for their isolation from the resulting biomass.
2. Materials and Methods
The objects of the study were the mother culture of the planktonic strain
Chlorella vulgaris BIN [
16,
17], the concentrate of the cell suspension of microalgae
C. vulgaris, and the extract of the total carotenoids isolated from the obtained biomass.
2.1. Cultivation
The cultivation of the microalgae was carried out according to the previously described method [
14] in a laboratory photobioreactor (PBR) with a volume of 10 dm
3. Combined additives were used to stimulate the biosynthesis of the carotenoids. As a stress inducer of carotenogenesis, a 3% solution of hydrogen peroxide (“Innovation”, Voronezh, Russia) was added to the nutrient medium. A solution of pyridoxine hydrochloride (“BioChemica”, Moscow, Russia) with an initial concentration of 50 mg/mL was used to prevent oxidative stress.
A culture medium should have the well-balanced content of macro- and microelements in order to cultivate microalgae [
18].
Table 1 shows the composition of the growing medium consisting of components used to prepare the culture media A and B, with the addition of macroelements after the mixing thereof.
The mother culture of
C. vulgaris BIN was removed from the refrigerating chamber (Melling YCD-519, Jackson, MS, USA) and introduced into the prepared PBR. The initial number of cells in the PBS was 8.0–8.5 million cells per mL. Then, the PBR was filled with the prepared culture medium. The stirring of the system during cultivation was carried out by bubbling with a **long AP-001 (Shantou, China) compressor at 1.5 L/min, and once a day with a mechanical stirrer for 15 min at 500 rpm (to prevent the formation of stagnant zones). The temperature range of the cultivation was 23 ± 3 °C. The illumination modes—“day–night” (16–8 h)—were maintained using LED TIS-15M1R lamps with a luminous flux of 3000 lumens [
19].
The change in the concentration of cells in the culture during the cultivation was monitored by the daily measurement of the optical density on a UV-1280 spectrophotometer (SHIMADZU, Duisburg, Germany) at a wavelength of 750 nm. The culture medium was used as a reference solution. The change in pH in the PBR was determined by using a Mini PH Meter (AQUA-LAB, Russia, Moscow). In order to study the morphological characteristics of the C. vulgaris cells, digital images of the “crushed drop” preparations were used with a magnification of at least 400 times using an IS-500 digital camera (LOMO JSC, St-Petersburg, Russia). The digital images were processed using the FOTO Microanalysis software (LOMO JSC, St-Petersburg, Russia) and Levenhuk (Levenhuk LabZZ manufacturer, China).
The additives were added to the PBR according to the following scheme: Day 1, without additives; day 2, 1 mL 3% hydrogen peroxide was added; day 3, 1 mL 3% hydrogen peroxide and 10 mL pyridoxine hydrochloride with a concentration of 50 mg/mL (the calculation of the additive amount was performed per 10 L of the culture medium). A cell suspension of microalgae without any additions was used as a control sample.
After the end of the cultivation cycle (4 days), the suspension was settled for 1 day. After settling, about 9 L of the suspension was poured through the lower drain hole of the PBS, leaving about 1 L PBS to start a new cultivation cycle. The specific growth rate of the biomass was determined by the following equation:
where
X0 and
X are the concentration of microalgae cells in the suspension before and after the cultivation, respectively, in million cells/mL; µ is the specific growth rate of biomass, day
−1; and
t is the cultivation time, in days.
2.2. Concentration of the Biomass
A solution 0.1 M NaOH was added to the cell suspension extracted from the PBR until a pH of 10–11 was reached. The formation of the flocks occurred within 2–3 h, after which the concentrate was separated from the culture medium by decantation, followed by the centrifugation of the sediment in a Thermo FS centrifuge Sorvall LYNX 6000 (Thermo Scientific, Waltham, MA, USA), at 6000 rpm/min for 5 min, followed by the decantation of the supernatant fluid.
2.3. Activation
The biomass was activated on a Silent Crusher M installation (IKA® Werke, T25 Basic, Heidolph, Germany) by high-speed ultra-homogenization (6000, 8000, 10,000, and 12,000 rpm for 5 min) in an ice bath, after the pre-mixing of the biomass with the liquid phase in a ratio of 1:5. A mixture of hexane and ethyl alcohol (95.6%) in a 2:1 ratio was used as the liquid phase. Pigments were then extracted from the obtained homogenate by ultrasonic extraction.
2.4. Extraction
In order to extract the pigments from the biomass, modified techniques were used [
20]. The pigments were extracted with a mixture of hexane and ethyl alcohol (95.6%) in a 2:1 ratio (
v/v) using an ultrasonic disperser (Scientz—IID, Zhejiang, Hangzhou, China). The ratio of the homogenate’s mass and the extraction mixture was 1:30 (by weight). The extraction process was carried out once. The separation of the pigment complex extract from the residual biomass was carried out using a Thermo FS centrifuge at 3300 rpm for 5 min. The resulting extract of the pigment complex was subjected to the fractionation.
2.5. Fractionation of the Pigments
The separation of the total carotenoids from the chlorophylls was carried out by saponification with KOH, followed by the separation of the fractions. The saponification process removed fatty acids, chlorophyll, and wax esters from the carotenoids.
KOH was added to the obtained extract in the amount of 0.05 g per 10 mL of the extract. The mixture was shaken for 10 min and then centrifuged using a Thermo FS centrifuge at 3300 rpm for 5 min. In this case, the carotenoid fraction (the upper phase in hexane) was colored yellow–orange, and the fraction of chlorophyllic acid salts (the lower phase in an aqueous solution of ethanol) was dark green. During the saponification, ethyl alcohol (95.6%) was added to the sample in order to prevent the precipitation of fatty acid salts.
The obtained extract of carotenoids in hexane was washed three times in a nitrogen atmosphere with cold distilled water in order to neutralize the pH, and was subjected to the concentration of the final volume of 0.5 mL using a LabTech Ev311Vac rotary evaporator (GWSI, Zhengzhou, China) in a vacuum (2 kPa) at a temperature of 45 °C. The resulting dry residue was analyzed for its carotenoid content.
2.6. Analysis of the Obtained Carotenoids
The carotenoid yield was determined spectrophotometrically on a UV-1280 (SHIMADZU, Duisburg, Germany) spectrophotometer in the spectral range of 350–700 nm. In order to determine the content of chlorophyll and carotenoids, the absorption bands of the pigments in the range of 470, 649, and 664 nm were used. The calculation of the chlorophyll and carotenoid concentration in the biomass of the microalgae was carried out according to the previously described procedure. Previously, we carried out studies and selected the conditions for the determination of carotenoids in extracts from
C. vulgaris biomass by reverse phase method HPLC [
21,
22].
The analysis of the individual carotenoids was performed using the HPLC technique. An autonomous modular device with a simple piston pump for HPLC Varian 03-919000-00 9010 Gradient HPLC Pump DT VAC Case Scratched, Discolored REF VA248 (USA) was used as the HPLC pump (USA). The eluent speed was 1 mL/min. In order to identify the carotenes and xanthophylls in the obtained extracts, standard samples of β-carotene (Merck, Germany) and fucoxanthin (Shanxi Fuhen Biotechnology Co., Ltd., ** pigment–protein complexes. For pigment–protein complexes, chlorophylls are stabilizers and an important cofactor due to their non-covalent interaction with the proteins. This means that, in order to maintain the stability of the newly formed pigment–protein complexes, it is necessary to immobilize a larger number of photosynthetic pigments, primarily chlorophylls, as well as their auxiliary pigment-antennae-carotenoids.
It is known that besides stimulating plastic processes, pyridoxine is able to partially neutralize the processes of oxidative stress. Farghl [
26] showed that pyridoxine significantly increased the activity of protective antioxidant enzymes in
C. vulgaris, such as peroxidase, superoxide dismutase, catalase and ascorbate peroxidase.
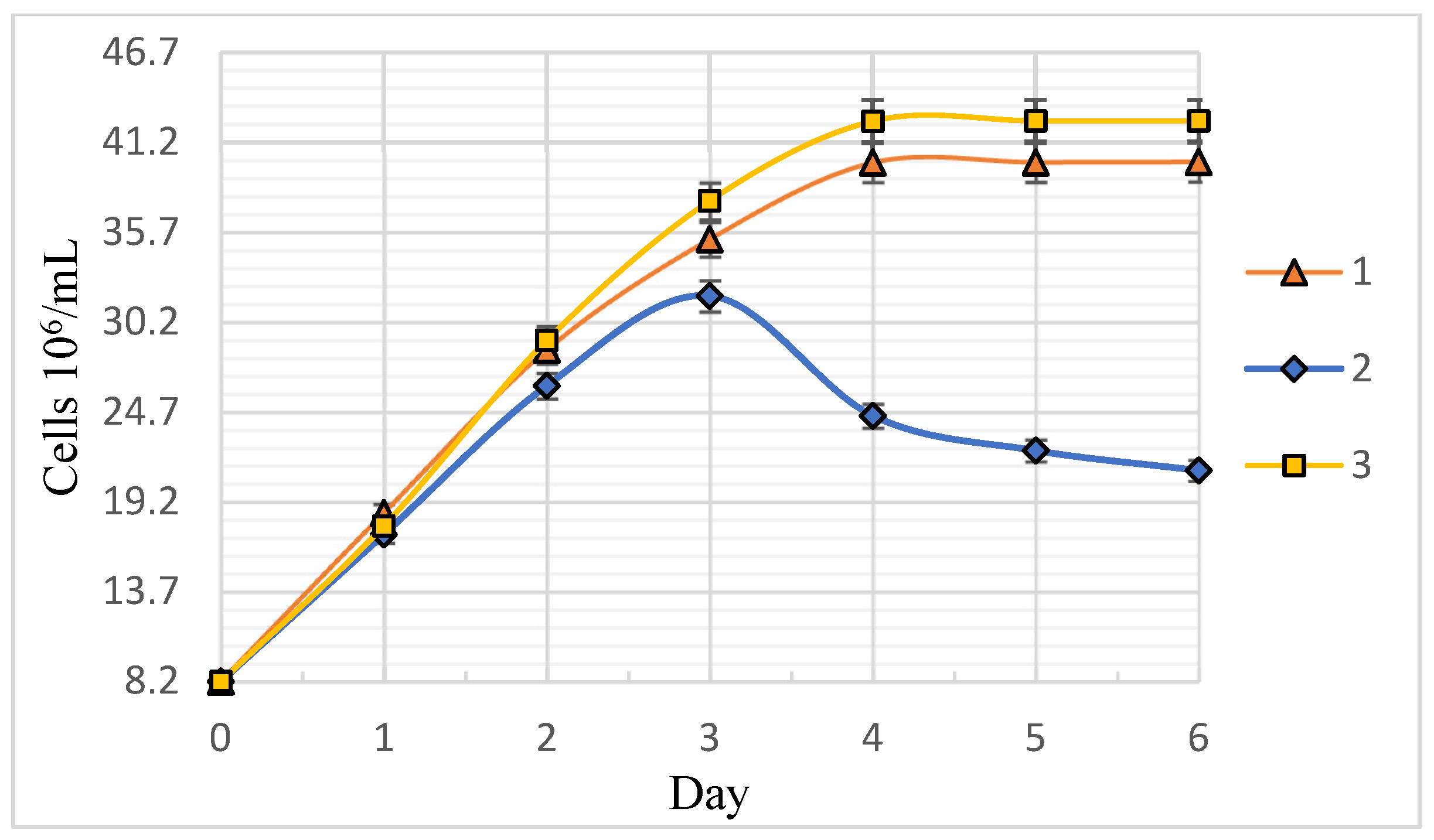
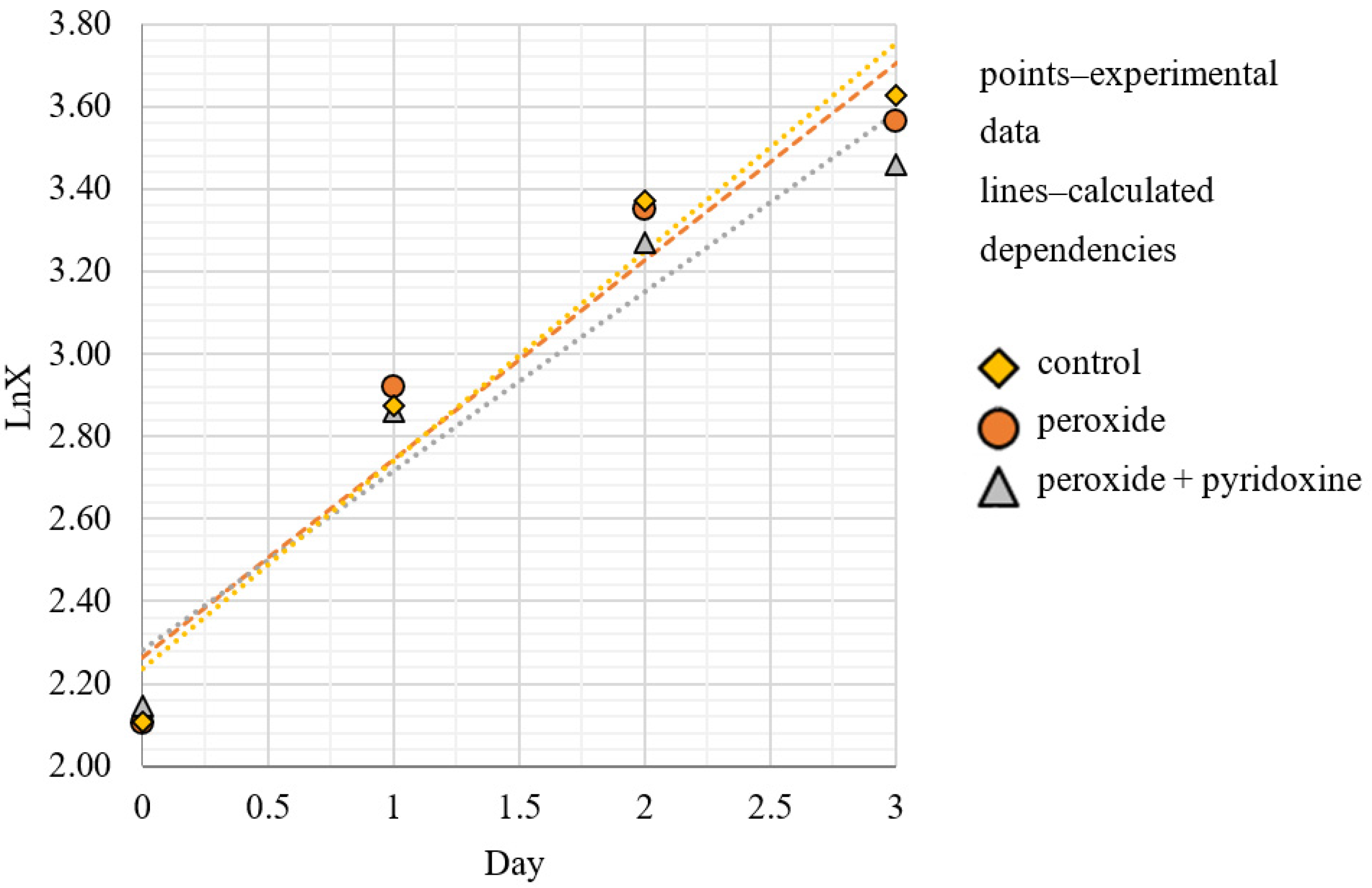
Figure 2 shows the exponential growth phase of
C. vulgaris BIN microalgae in semi-logarithmic coordinates. The approximation of the exponential growth phase is a straight line with the slope tangent corresponding to the value of the population-specific growth rate μ, in day
−1. The specific growth rate varied from 22 to 25 days
−1. It was noted that the cell fluid of sample 3 had a more saturated dark green color, indicating the increased synthesis of photosynthetic pigments.
Figure 3 shows the effects of the additives on the pH dynamics in the process of
C. vulgaris cultivation. The relationship between the cultivation curves and pH dynamics is noted. According to Farghl [
26], it is connected to the metabolism of the cells and their photosynthetic activity. The work [
27] reports that medium alkalization can occur due to algae absorbing the nitrate form of nitrogen due to the export of ammonium ions’ excess during photorespiration and other metabolic processes. The viability indicator of
C. vulgaris microalgae is the increase of the medium’s acidity agreeing well with the dynamics of the biomass accumulation shown in
Figure 1.
3.2. Spectral Characteristics and Carotenoid Content of C. vulgaris BIN Biomass Extracts
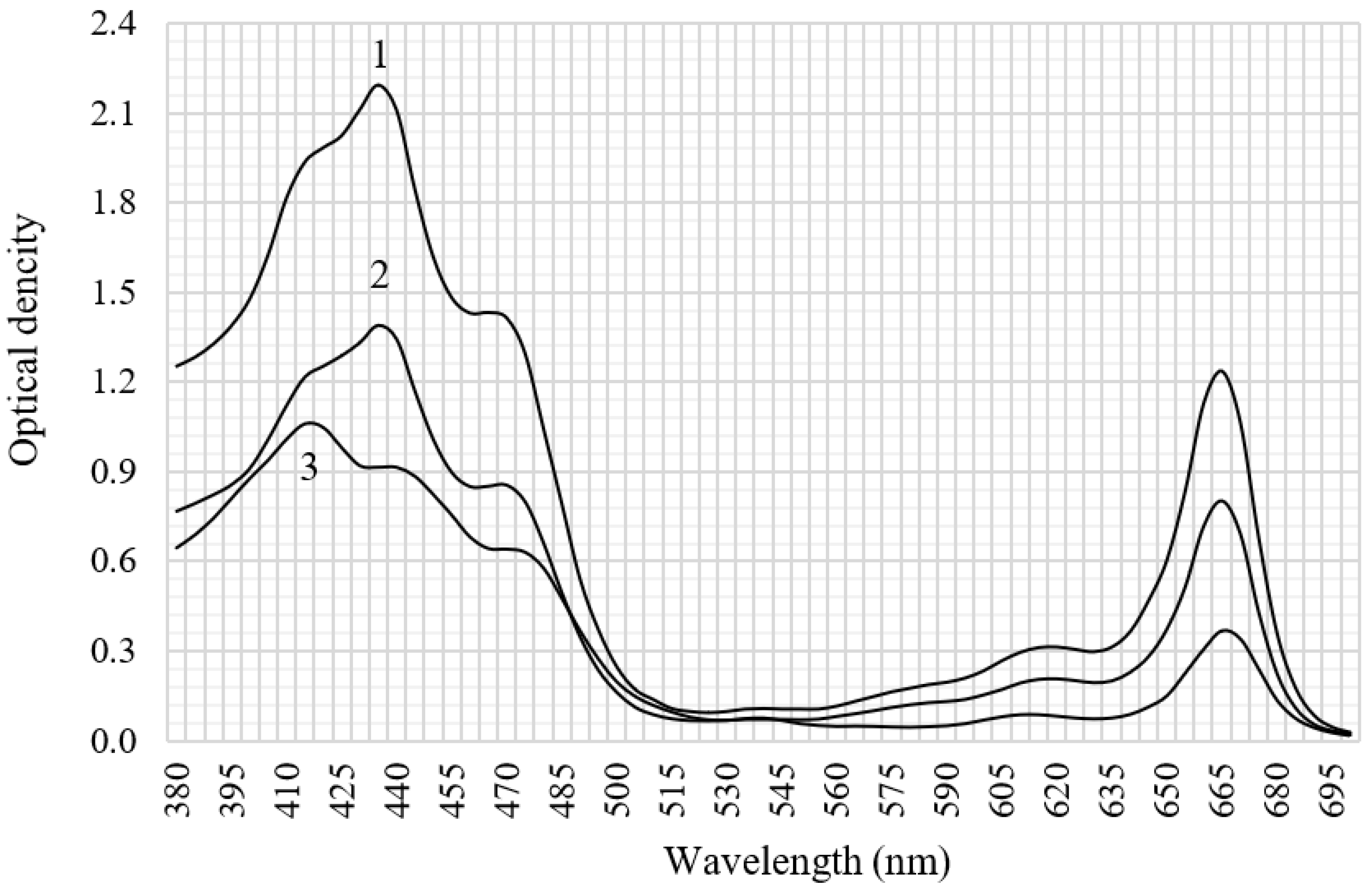
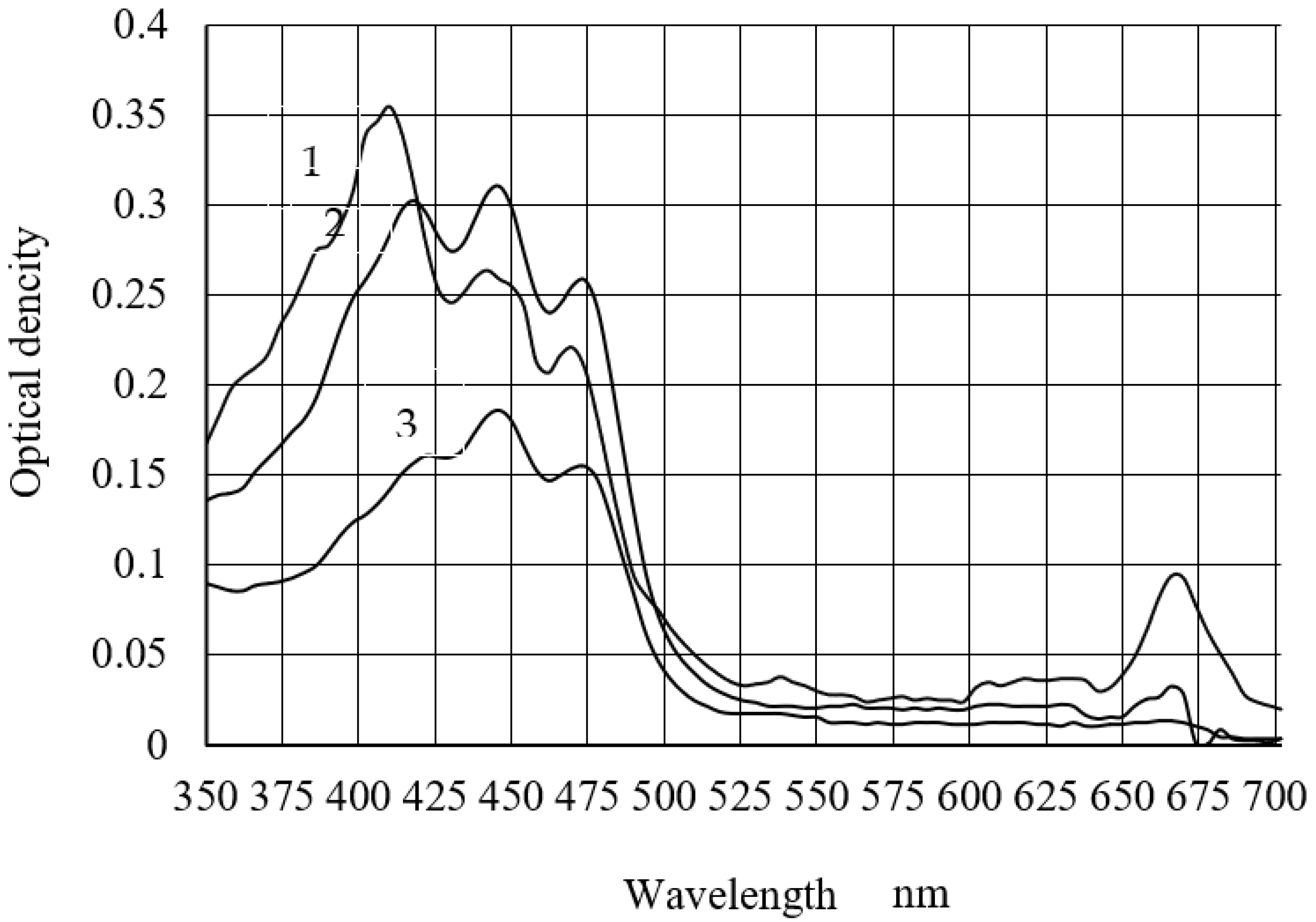
The cultivation factors’ influence on the spectral profiles of pigment extracts from the microalgae is shown in
Figure 4. Two absorption bands were noted in the blue–violet (380–500 nm) and in the red (600–680 nm) spectrum range. The arm (420 nm) and the absorption maximum (664 nm) correspond to chlorophyll a. The carotenoid absorption band is in the spectral range of 420–480 nm, with absorption maxima of 440 and 470 nm [
28,
29]. This allows us to talk about the predominance of carotenoids in the pigments of sample 3.
Figure 5 shows the composition of photosynthetic pigments in the obtained samples of
C. vulgaris biomass. The highest content of carotenoids was observed in the biomass sample with H
2O
2 and pyridoxine additives, while the content of carotenoids was about 1.8 times higher than that in the control sample. A biomass sample with H
2O
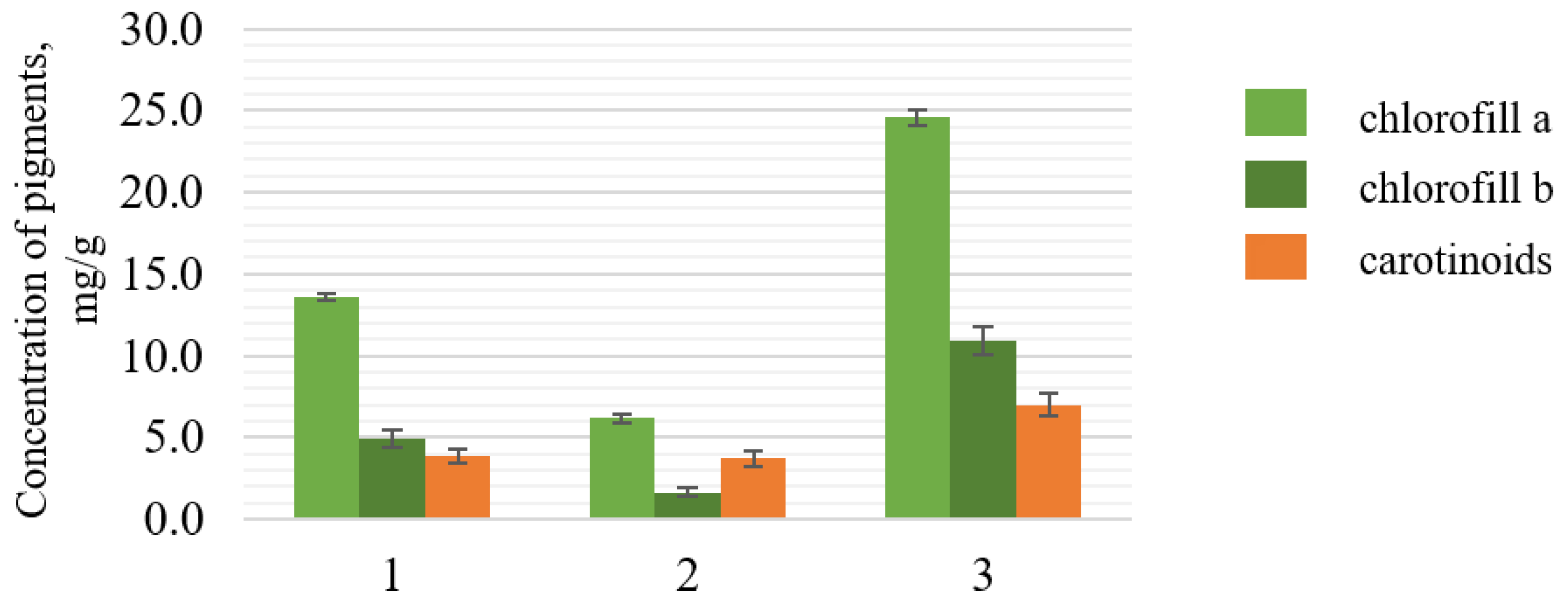
2 showed a 0.2 mg/g reduction in carotenoids compared to the control sample. The contents of chlorophyll a and b in samples show the same logic.
The highest content of carotenoids was observed in the extracted biomass with added H2O2 and pyridoxine; the content of carotenoids was about 1.8 times higher than that in the control sample.
3.3. Influence of Activated C. vulgaris BIN Biomass on the Yield of Carotenoids in the Extracts
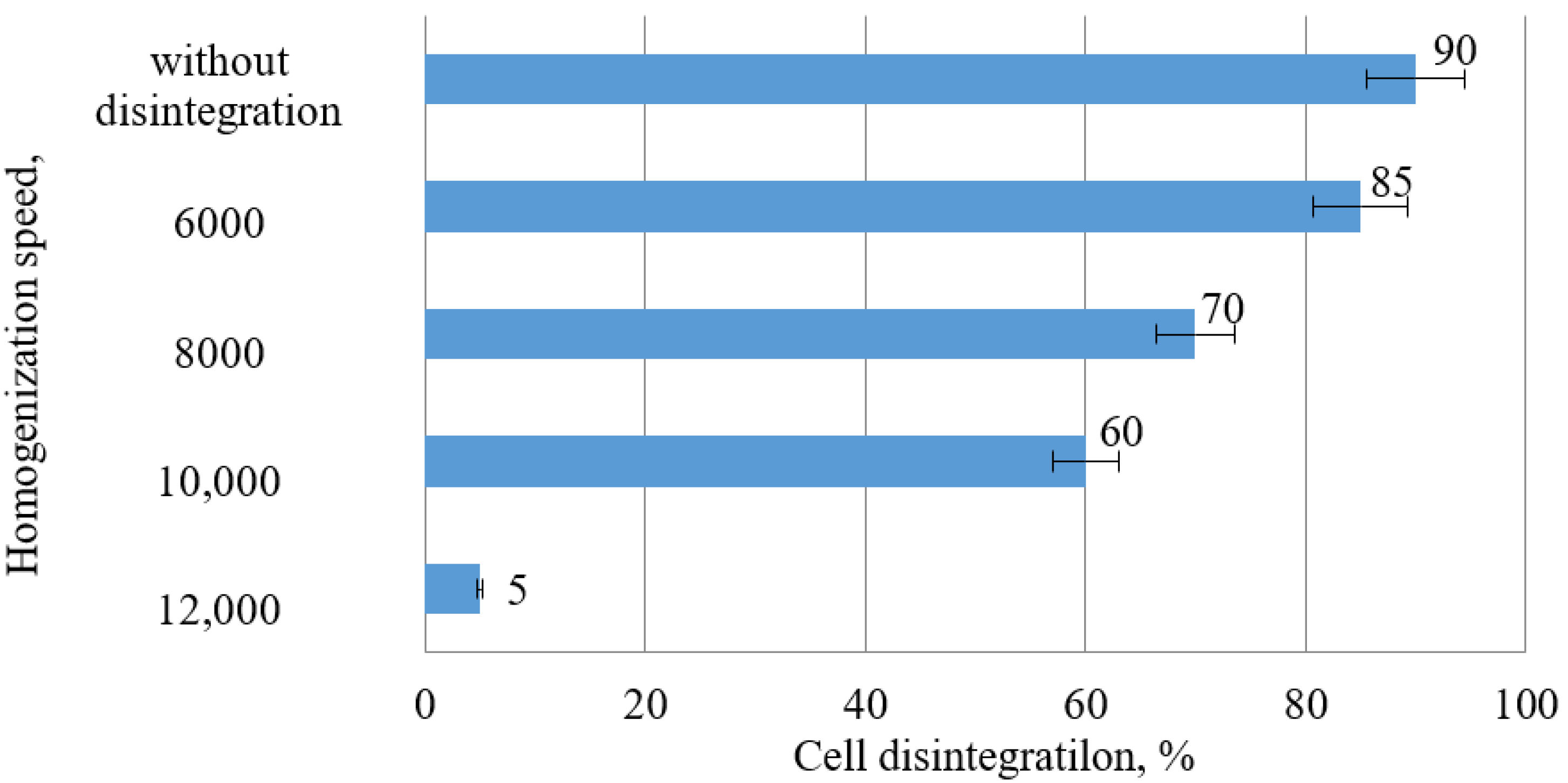
The influence of high-speed homogenization on the microalgae activation rate, evaluated with cell microscopy, is shown in
Figure 6.
Table 2 shows the effect of the rate of homogenization on the yield of carotenoids in the extracts.
It was found that an increased rotor speed of more than 6000 rpm leads to a significant increase of destroyed cells, causing the mixing of plastid pigments with oxidases, as well as the rapid oxidation thereof. This was proven by the decreased yield of carotenoids in the extracts. Thus, the most optimal mode of cell membrane activation with preserved cell integrity and shape is the rotation speed of 6000 rpm.
3.4. Extraction and Fractionation of the Pigments
We proposed to use a mixture of hexane and ethanol alcohol in a ratio of 2:1 as an extracting agent, which resulted in the high yield of the pigment complex.
The saponification step to separate chlorophylline acids is the key to obtain the carotenoid fraction. Saponification at low KOH concentrations (0.01 and 0.2 M) gives the best results for astaxanthin extraction.
Long-term saponification with relatively high KOH concentrations (0.2 and 0.5 M) gives the best results for xanthophyll extraction.
The research of the KOH concentration’s effect on the spectral profile of
C. vulgaris extracted pigments is shown in
Figure 7. It was found that an alkali concentration of 0.2 M is most appropriate, and that it allows us to obtain the highest concentrations of the determined substances.
The alkali-free extract of carotenoids in hexane had a pH level of 5.5. Shivalkar and Prabha [
30] reported that the major stability of carotenoids is revealed in the pH range from 6 to 7. Thus, it is required that we remove excess alkali after the carotenoids’ saponification in order to obtain neutral pH values. The required pH values are reached after the third stage of washing with oxygen-free distilled water in an inert atmosphere.
It was found that the amount of impurities (carotenoid ethers and chlorophyll derivatives) in the extract was reduced. The carotenoid loss amounted to an average of 6%. The spectral profile of the carotenoids showed a distinctive “corona”, with three peaks at 420, 445 and 472 nm. A complex of peaks similar to those obtained was described by the author for neurosporene (414, 439, 467 nm), and by the authors for α-carotene (423, 444, 473 nm) and β-carotene (425, 450, 478 nm) [
31].
The carotenoid pigments’ composition is analyzed in
Table 3.
The obtained sample of carotenoids had a bright orange color, and the content of the main substance was 52 ± 3 mg/mL. The carotenoids of C. vulgaris microalgae are represented by lutein (about 45%), fucoxanthin (about 12%) and β-carotene (4.3%). The total carotenoids were resuspended in soybean (castor) oil in order to obtain the oil concentrate that we propose to use as a dietary supplement in the future.
4. Conclusions
The application of carotenoids to produce functional products is not easy due to the low bioavailability of the pigments in their native form. Therefore, the search for additional sources of carotenoids is of paramount importance. Innovative food and additives based on hydrobionts with a high content of carotenoids are promising in this sphere.
This study showed that microalgae can be successfully used as a source of carotenoids. In order to prepare the biomass for the further extraction of the necessary pigments, the directed cultivation of C. vulgaris BIN was used. The pigments were efficiently extracted using ultrasound. The carotenoid sample obtained after vacuum thickening contained the main substance in the amount of 52 ± 3 mg/mL. The product’s composition was determined: lutein, 45%; fucoxanthin, 12%; and β-carotene, 4.3%.
The obtained sample of carotenoids was further prepared in order to develop the functional product enriched with bioactive compounds. The carotenoid sample was re-dissolved in castor oil to produce a microencapsulated product for targeted delivery to the lower gastrointestinal tract. This potential application is of high importance because of the growing demand for functional products contributing to a healthy lifestyle.


 ,
, 








{kind=link}
{kind=link}
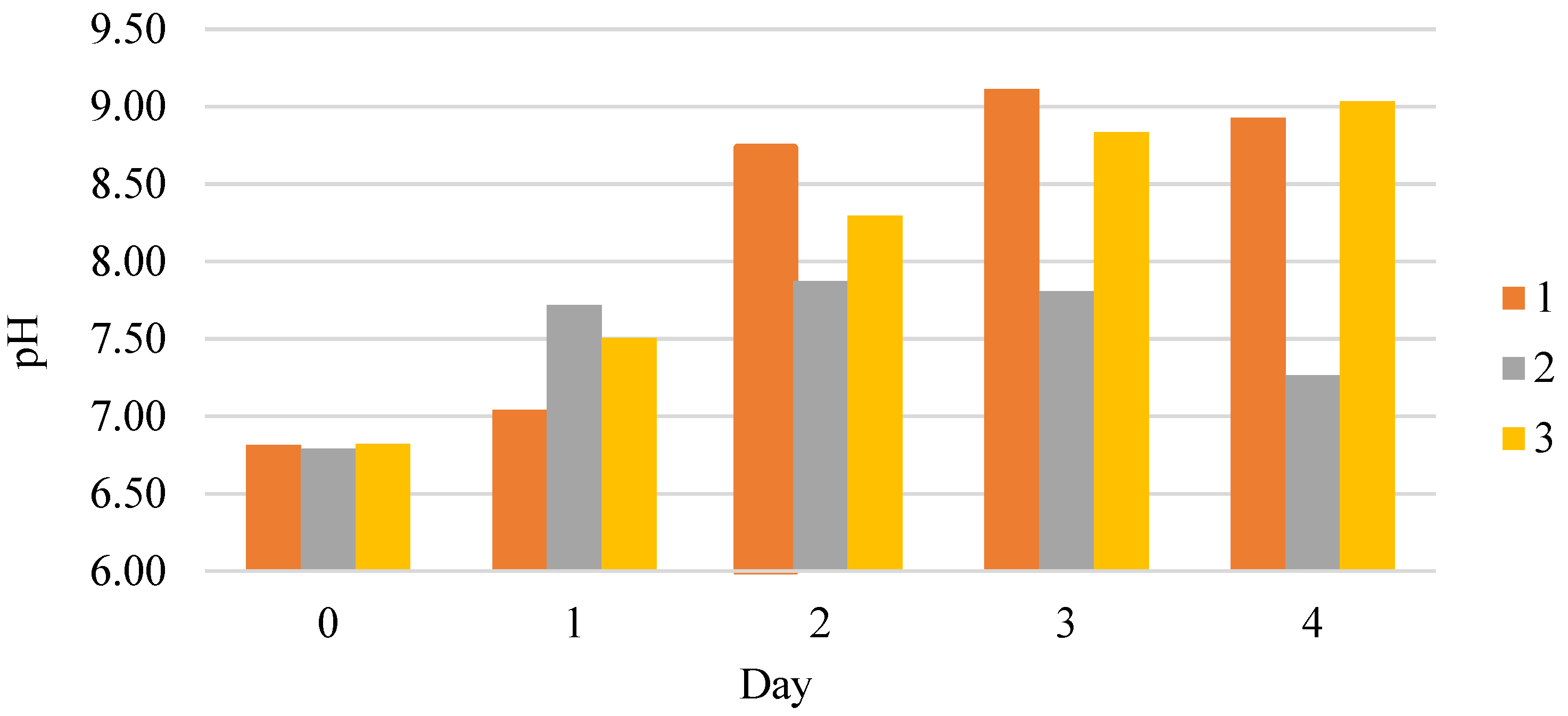{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}