1. Introduction
Angiostrongyliasis is a parasitic zoonosis disease caused by the nematode species
Angiostrongylus cantonensis and
A. costaricensis. It is endemic in the Southeast Asia-Pacific Basin region and the American continent (the Caribbean and South America) [
1,
2,
3]. Still, the parasite has also been found in Australia, some areas of Africa, Hawaii, Louisiana, and more recently in Europe [
4]; it is reported that at least 2800 cases have been recorded globally [
5].
A. costaricensis causes an abdominal disease, while the
A. cantonensis an eosinophilic meningoencephalitis. The first was described in Costa Rica in 1952 [
6,
7,
8] and the second infecting pulmonary arteries and hearts of domestic rats in Guangzhou (Canton), China, by Chen in 1935 [
9]. In both cases, rats are the primary hosts, and snails are the intermediate hosts [
10,
11].
Diagnosis is sought via clinical criteria, including the presence of cerebrospinal fluid eosinophils and a history of exposure to
A. cantonensis larvae (third-larvae, L3), e.g., from raw freshwater snails (Pila or Pomacea snails) or contaminated vegetables [
12,
13]. Conversely, the diagnosis based on clinical presentation is difficult because the symptoms are often confused with those produced by other parasitic diseases such as paragonimiasis, gnathostomiasis, and cysticercosis [
14]. A definitive diagnosis is confirmed by discovering the worms in cerebrospinal fluid after a lumbar puncture or in the eyes during surgery, but direct detection of the parasite in patients is rare [
2,
15].
Over the past decades, many non-particulate proteins (adult worms, brain-stage larvae, or excretory-secretory products) have been evaluated as potential targets for the development of immunology to support the clinical diagnosis [
16,
17,
18,
19]. The enzyme-linked immunosorbent assay (ELISA) [
20,
21,
22,
23], dot blot [
16,
23,
24,
25], and immunoblot [
26] with extract of the parasite [
27,
28] or using proteins isolated by electroelution [
27] have been most widely applied. More recently, recombinant proteins derived from these previously identified proteins were also applied to a lateral-flow immunochromato graphic test assay [
29,
30,
31]. However, in the totality of these works, antigens highly conserved in nematodes and other correlated genes were used, which may suggest cross-reactive epitopes [
32,
33], and their specificity and validation have not yet been present bee confirmed in studies involving multi diseases patients sera.
Next-generation metagenomic sequencing for diagnosing angiostrongyliasis of the central nervous system is being validated as a possible diagnostic methodology [
34]. However, the fact that the number of
A. cantonensis DNA readings has fluctuated with the opening pressure of the CSF and clinical manifestations calls into question the validity of the methodology for laboratory diagnosis [
35].
Therefore, given the difficulties of applying molecular tools, the development of immunological methods continues to be a viable and necessary alternative for the quick and accurate diagnosis of angiostrongyliasis. Consequently, initial protein characterization is crucial in reaching an excellent immunological diagnostic or therapeutic target [
25,
36].
Proteases are critical elements of several biological processes in living cells and, therefore, represent one of the most prominent families of biopharmaceutical targets. Among this group of proteins, the traditional family of aspartyl proteases stands out, and within these, a new group of polytopic membrane proteins called presenilin (PS) presents extremely divergent sequences outside of some well-conserved motifs. This group of proteins has hardly been studied for drug development [
37] but may also serve as antigens [
38,
39,
40], and the discovery of new families of membrane-associated aspartyl proteases is growing in eukaryotic cells [
41,
42,
43]. Notably, it has been estimated that proteases account for 5 to 10% of all drug targets studied for therapeutic development [
10].
Conversely, to inhibit and block the activity of pathogenic proteases or to identify specific epitopes with the desired selectivity, monoclonal antibodies (mAbs) have shown great promise as research tools, therapeutic agents, and diagnostics. However, identifying mAbs with inhibitory functions is challenging because current antibody discovery methods rely on binding rather than inhibition. Consequently, identifying epitopes located in strategic domains of these proteins through peptide microarray presents a faster and more efficient alternative to identify these determinants and then produce the desired antibodies.
Therefore, in this study, we describe the partial biochemical and immunological characterization of a putative presenilin protein of A. cantonensis and its epitopes in develo** an ELISA test with high specificity and sensitivity for the diagnosis of the angiostrongyliasis.
2. Materials and Methods
2.1. Rat Infected Sera and Ethics Statement
Sera samples from 16 infected rats in the acute phase of angiostrongyliasis were obtained from the Laboratory of Biology and Parasitology of Wild Mammals Reservoirs (Oswaldo Cruz Institute, FIOCRUZ, Brazil). Another group included 25 uninfected rat sera.
2.2. Synthesis of the Cellulose-Membrane-Bound Peptide Array
A library of eight one, 15-mer peptides were designed to represent a consecutive overlap** coverage offset by nine amino acids across the entire coding region (415 aa) of the putative aspartyl protease (C7BVX5_ANGCA, UNIPROT) of
A. cantonensis. The peptides were automatically prepared onto amino-PEG
500-UC540 cellulose membranes (Intavis Bioanalytical Instruments, Köln, Germany) as previously described [
44] using an Auto-Spot Robot ASP-222 (Intavis Bioanalytical Instruments AG, Köln, Germany) and the F-moc strategy. In brief, coupling reactions were followed by acetylation with acetic anhydride (4%,
v/
v) in
N,
N-dimethylformamide to render peptides unreactive during the subsequent steps. After acetylation, F-moc protective groups were removed by adding piperidine to render nascent peptides reactive. The remaining amino acids were added by this same process of coupling, blocking, and deprotection until the expected desired peptide was generated. After adding the last amino acid in the peptide, the amino acid side chains were deprotected using a solution of dichloromethane–trifluoracetic acid–tri isobutyl silane (1:1:0.05,
v/
v/
v) and washed with methanol. Membranes containing the synthetic peptides were either probed immediately or stored at −20 °C until required. Negative controls [without peptide; IHLVNNESSEVIV HK (
Clostridium tetani) precursor peptide] and positive controls were included in each assay.
2.3. Screening of SPOT Membranes
SPOT membranes were washed with TBS (50 mM Tris-buffer saline, pH 7.0) and blocked with TBS-CT (Tris-buffer saline, 3% casein, 0.1% Tween 20, pH 7.0) at room temperature under agitation or overnight at 4 °C. After extensive washing with TBS-T (Tris-buffer saline, 0.1% Tween 20, pH 7.0), the membrane presenting the peptide library was incubated for two hours with a pool of patients sera (1:100) in TBS-CT and then washed again with TBS-T. Afterward, the membrane was incubated with alkaline phosphatase-labeled goat anti-rat IgG (1:5000 in TBS-CT; KPL, Gaithersburg, MD, USA) for one hour, and then washed with TBS-T and CBS (50 mM citrate-buffer saline, pH 7.0). Chemiluminenscente CDP-Star® Substrate (0.25 mM) with Nitro-Block-II™ Enhancer (Applied Biosystems, Waltham, MA, USA) was added to complete the reaction.
2.4. Scanning and Measurement of Spot Signal Intensities
As described previously, chemiluminescent signals were detected on an Odyssey FC (LI-COR Bioscience, Lincoln, NE, USA) [
45]. Briefly, a digital image file was generated at a resolution of 5 MP, and the signal intensities were quantified using the TotalLab TL100 (v. 2009, Nonlinear Dynamics, Newcastle, Tyne, UK) software. The signal intensity (SI) used as a background was a set of negative controls spotted in each membrane.
2.5. Synthesis of Peptides and Preparation of the MAPs
Nine single
A. cantonensis peptides (PSAg1-PSAg3, PSAg6-PSAg9, PSAg11, and PSAg12;
Table 1) were synthesized by the F-moc strategy in a synthesizer machine (MultiPep-1 CEM, Corp, Charlotte, NC, USA). For the preparation of the dendrimer multi-antigen peptides (MAP4), the same synthesis protocol was used in the solid phase and the tetrameric F-moc-Lys2-Lys-β-Ala Wang resin, as described previously [
46]. The constructs were prepared in the automated MultiPep-1 peptide synthesizer, and the side chains of tetrafunctional F-moc-amino acids were protected with TFA-labile protecting groups as required. Residues corresponding to the monovalent (‘tail’) part of the construct, up to the first (bis-Fmoc) Lys residue initiating the dendrimer structure, were incorporated via single couplings. Once sequence assembly was completed, the F-moc groups were removed, and the peptide-resin was cleaved and fully deprotected with TFA/H2O/EDT/TIS (94/2.5/2.5/1.0
v/
v, 90 min). The peptide was precipitated by adding chilled diethyl ether, centrifuged for 3 × 10 min at 4 °C, and the pellet was taken up in aqueous AcOH (10%
v/
v), dried, and stored as a lyophilized powder. When necessary, the MAP was dissolved in water, centrifuged (10,000 g, 60 min, 15 °C) and the supernatant filtered on a centricon 10 filter. The single peptides were used without previous purification, but their identity was checked by MS (MALDI-TOF or electrospray).
2.6. ELISA-Peptide
The ELISA was realized, as described previously [
47], with minor modifications. Briefly, ELISA plates (Immulon 4HB; Corning, NY, USA) were coated with 80 µL (50 µg) of each peptide prepared on coating buffer (Na
2CO
3–NaHCO
3, pH 9.6) overnight at 4 °C. After each incubation step, the plates were washed three times using a PBS-T washing buffer (PBS with 0.1% Tween 20 adjusted to pH 7.2) and blocked (200 µL) with 2.5% BSA and incubated 2 h at 37 °C. The rat’s sera were diluted (1:150) in coating buffer, and 100 µL were applied onto immunosorbent plates and incubated for 2 h at 37 °C. Following several washes with PBS-T, the plates were incubated with 100 µL goat anti-rat IgG conjugated to HRP (1:5000 dilutions at blocking buffer; KPL, Gaithersburg, MD, USA) for 2 h. The plates were developed with p-nitrophenyl phosphate (p-NPP) as a substrate (Sigma-Aldrich, St. Louis, MO, USA). The absorbance values at 405 nm were read using a FlexStation 3 Microplate Reader (Molecular Devices, Sunnyvale, CA, USA), and the reactivity index was defined as O.D.
450 value of Target—O.D.
450 values of cut-off.
2.7. Database Searches, Computational, and Phylogeny Studies
To curate a family of 10 target sequences for C7BVX5-1, a set of aspartic protease sequences were retrieved from the Uniprot database (available online:
http://www.uniprot.org/ accessed on 20 March 2021) using the following criteria: An EC designation of 3.4.23 for aspartic endopeptidases; a length between 300 to 800, and a reviewed annotation. Another set of sequences homologous to other proteins, which have been cited by Blast, was added after a delta-blast of the Refseq database with coverage greater than 80% of their sequence-issue and identity over 10–30%. From this set, an alignment was performed on the Mat server with blossum80 and a gap penalty of 2.5, followed by clustering to eliminate the excessive gaps with the method “minimum linkage.”
The potential transmembrane (TM) domains (TMD) of the
A. cantonensis enzyme were analyzed by three different predictions programs; the TMpred [
48], the TopCons [
49], and MemConP [
50]. The model most consistent with the identified epitopes and motifs were obtained using TopCons, which used an algorithm based on the statistical analysis of base. The prediction was made using a combination of several weight-matrices for scoring. Secondary structure predictions were obtained from PSIPRED (Available online:
http://bioinf. cs.ucl.ac.uk/psipred/ accessed on 15 March 2021) and CDM (available online:
http://gor.bb.iastate.edu/cdm/ accessed on 15 March 2021) servers. The tertiary structure prediction was performed on the LOMETS server (available online:
http://zhanglab.ccmb.med.umich.edu/LOMETS/ accessed on 15 March 2021) and I-TASSER (Available online:
https://zhanglab.ccmb.med.umich.edu/I-TASSER accessed on 15 November 2020). Finally, a network analysis was performed using STRING tutorial and the Cytoscape platform version 3.7.2; (available online:
https://apps.cytoscape.org/apps/stringapp accessed on 21 October 2021) [
51].
2.8. Ethics Statement
The study was approved by the FIOCRUZ (IOC-CAAE: 52892216.8.0000.5248 and 1.896.362) study center ethics committee and conducted following all applicable Good Clinical Practice (GCP) regulatory requirements, including the Declaration of Helsinki.
2.9. Statistical Analysis
Statistical analysis was performed using GraphPad Prism version 5.0. The statistical differences using a t-test were considered if the p-value ≤ 0.05.
4. Discussion
Our group has been working for some years on the characterization of GSC proteins from lower eukaryotes [
53,
54] and we observed that this group of proteins has a very low homology (~20–30%) to each other, a fact that has made their identification difficult in genomics and proteomics [
55,
56] projects. However, on the other hand, this information points out that these proteins are more likely to have specific epitopes under the immunological point of view.
Therefore, to overcome the difficulty of the diagnostic specificity of angiostrongyliasis, we characterize a protein from A. cantonensis described in the database as a putative presenilin protein using bioinformatics, biochemical and immunological tools. This first characterization of an atypical transmembrane aspartyl protease from A. cantonensis.
Intramembrane proteases catalyze the unusual cleavage of peptide bonds in the plane of biological membranes. They are categorized according to their active site. For example, the MEROPS database (MEROPS database. Available online:
https://www.ebi.ac.uk/merops/ accessed on 18 November 2021) peptidase family, A22, contains membrane-inserted endopeptidases and aspartates residues in the active area are characteristics of peptidases of family A22. The GxGD aspartyl proteases comprise the classical human PS1 and PS2, the signal peptide peptidase (SPP), and SPP-like (SPPL) proteases [
57,
58] and five homologous presenilin proteins,
Presenilin is the catalytic portion of the gamma–secretase complex (GSC) along with other proteins, Anterior pharynx defective 1 (Aph1), Nicastrin, and presenilin enhancer 2 (Pen2), involved in complex stabilization and substrate binding. Most of the presenilins characterized until now are composed of 9-pass TM domains that undergo an endoproteolytic cleavage in the intracellular loop between TM6 and TM7, producing N-terminal and C-terminal fragments [
59,
60].
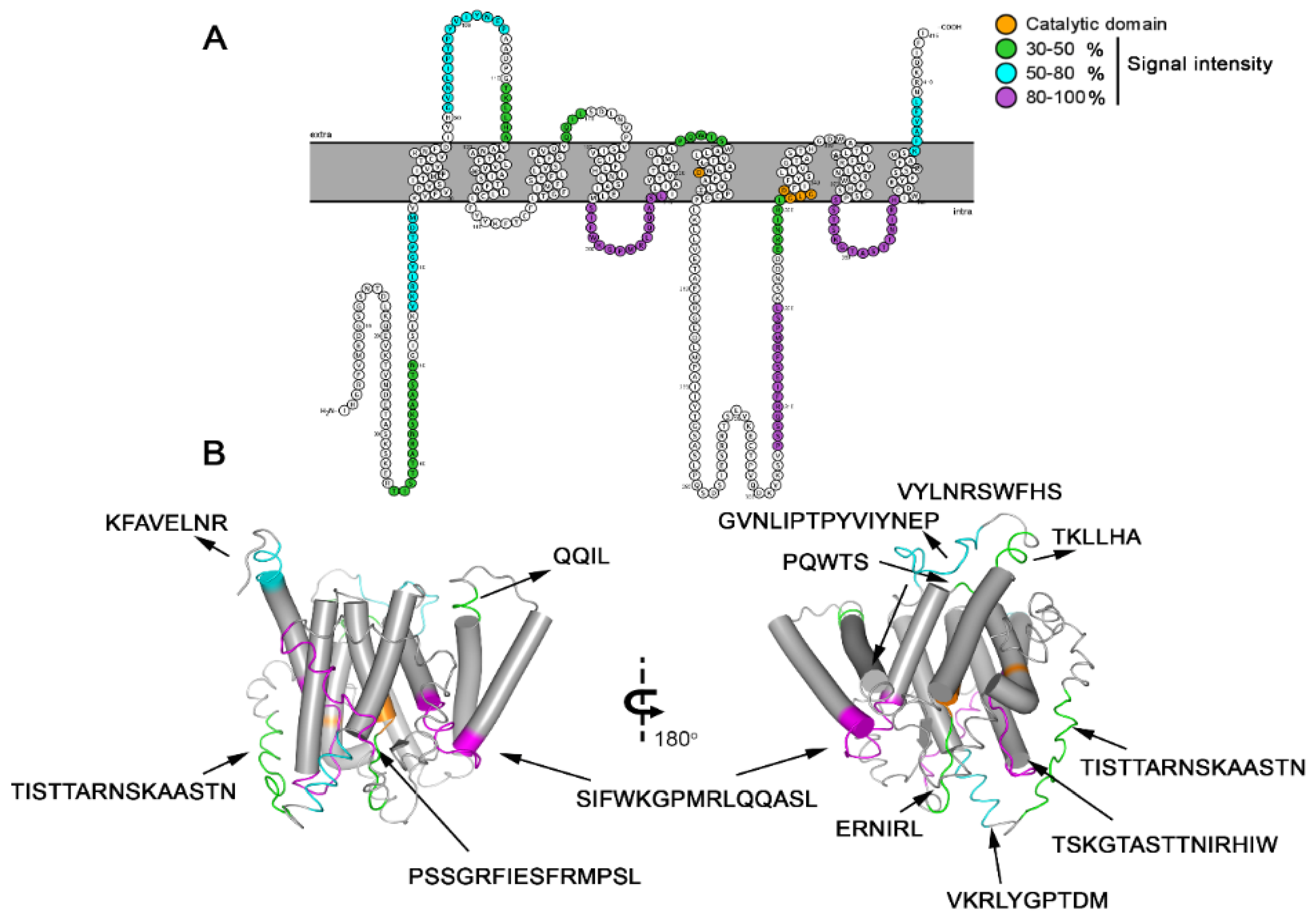
The predicted model for Ag-PS protein also presented 9 TM domains. Its topology is corroborated by the experiments of epitope identification (outside and inside) and contemplates the catalytic domain entailed of the conserved aspartates motifs XD (aa 241) and GLGD (aa 332-335), between TM6 and TM7, as occur in the human orthologue (YD; 192–194 and GLGD; 287–290) protein (
Figure 2)
. The presence of these critical motifs was also confirmed to be present in other worms such as
Caenorhabditis elegans (P52166),
Angiostrongylus costaricensis (A0A0R3PEK8), and
Schistosoma mansoni (C0SKM6) (
Figure 2).
However, the AgPS structure does not present other significant functional motifs such as the [GxxxG (GVKLG), SxxxGxxxxA (SVLVGKASA)] and the PALP. The first two motifs should be located at TMD8 of other eukaryotic proteins closely to the catalytic Asp and contribute to the helix packing. The PALP in the TMD9 stabilizes the GSC and characterizes the A22 family. This function has been demonstrated in studies performing mutations in the first Proline of PALP with leucine (P414L) which abrogated PS activity in
Drosophila melanogaster and
C. elegans [
61].
Previously, a 46 kDa soluble aspartyl protease (GenBank accession no. NP 872129) from
A. cantonensis was cloned and expressed and present in different infective levels larvae, young adults, and adult worms. Its primary function has been associated with exogenous hemoglobin degradation [
62].
In this work, we describe, for the first time, the presence of an atypical presenilin-like aspartyl protease (AgPSat) in
A. cantonensis. This nomenclature was attributed to proteins from different organisms that have high similarity with PSs but do not present certain domains characteristic of the PS family [
62]. These non-catalytic activities may represent an ancestral PS function, described in non-mammalian animals, plants, and less complex eukaryotic organisms [
63].
This sub-branch also possesses two conserved aspartic acid residues within adjacent predicted TM segments. However, the branch of presenilin homologous (PSH) proteins retains the conserved first Proline and the PALP motif but lacks several other motifs present in the A22 family [
64,
65]. Contrasting in the Ag-PS, the PALP motif is absent, but the protein has a proline located in the C-terminal fraction (aa 373). Interestingly, phylogenetic close related worms such as
Haemonchus contortus (
Figure 4) have the PALP motif. These collective data suggested that AgPS is an atypical aspartyl protease, with catalytic and non-catalytic functions essential for worm homeostasis [
66,
67,
68,
69,
70,
71].
The tissue and cellular location of this critical proteolytic enzyme and its cellular function need to be demonstrated.
To characterize this TM protein immunologically, a microarray of peptides was initially synthesized using the Spot-synthesis technique to identify linear B-cell epitopes in the putative PS of
A. cantonesis that are recognized by infected rats IgG antibodies. Twelve epitopes were readily defined (
Table 1), confirming the predicted AgPSat protein’s likelihood to be expressed. In addition, the distribution of the peptides corroborates the topological model proposed for the AgPSat.
All peptides sequences were apparently specifically present in the
A. cantonensis protein from the determinant profiles. Thus, initially, the immunogenicity of the 12 peptides was demonstrated by an ELISA-peptide assay using a pool of infected rats sera (
Figure 4). An ELISA assay was developed using one of the peptides that highly showed sensitivity and specificity of MAP4, displaying its potential role to overcome the limitations of
A. cantonensis diagnostics.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}