3.1. Agar-Well Diffusion
Initial screening of the antimicrobial activity of the investigated agents was studied against the four microorganisms using the agar-well diffusion test, which showed the presence and absence of ZoI (
Table 1). All agents revealed different degrees of antibacterial activity against the tested bacteria. The antibacterial action was classified following the Rota et al. scale, which reports weak activity with a ZoI (halo) ≤ 12 mm, moderate activity with a ZoI ranging between >12 and <20 mm, and strong activity with a ZoI zone ≥ 20 mm [
35,
36]. AgNPs display a low-activity ZoI, particularly against Gram-negative bacteria. It has been reported that metal nanoparticles tend to form agglomerates when in colloidal dispersions, which reduces their diffusivity, limiting the contact with the bacteria [
37]. Here, the presence of agglomerates is evident (
Figure 1), supporting this statement. The agar tortuosity may also influence this phenomenon by hindering the AgNPs permeation through the culture media. Vancomycin antibiotic action occurs at the bacterial cell wall through the disruption of the synthesis of its major constituent, peptidoglycan. Vancomycin binds to the terminal carboxyl group of D-alanyl-D-alanine within nascent peptidoglycans of the cell wall, preventing the formation of lipid II, a key “shuttle carrier” of the peptidoglycan monomers [
38,
39]. This molecule forms a series of hydrogen bonds with the peptide backbone blocking its processing [
40,
41,
42]. It is true that Gram-negative bacteria outer cell wall is fairly impermeable to large glycopeptide molecules such as vancomycin [
2]. Therefore, vancomycin was reported as weak against the Gram-negative bacteria but strong against the Gram-positive bacteria. As such, the absence of the outer membrane and periplasm in the Gram-positive bacteria allows a higher permeability of vancomycin through its cellular wall, which possess a hydrophilic porous structure [
43,
44,
45].
AMPs LL37 and pexiganan are well-known effective antimicrobial agents. However, data from
Table 1 revealed a weak to moderate ZoI against the selected bacteria, respectively. The action of LL37 is suspected to have been compromised by the presence of salts within the solvent, PBS (recommended by the manufacturer), as the LL37 structure varies with the ionic charge of the solvents, possibly reducing its agar diffusion capacity [
46]. On its turn, pexiganan action was considerably superior to that of LL37, particularly against
S. epidermidis and
P. aeruginosa. These findings are consistent with the literature [
18].
The EOs displayed a variable degree of antibacterial activity against the tested bacteria. Interestingly, they were all found to be more effective against the
S. aureus bacteria, with a moderate (NO) to strong (TTO and CLO) activity. These results were closely followed by the
S. epidermidis and
E. coli bacteria.
P. aeruginosa was the least susceptible to the EOs action, with weak (NO) to moderate (TTO and CLO) ZoIs being formed. Once again, this can be explained by the differences in cell wall structure between Gram-positive and Gram-negative bacteria, with the latter being capable of restricting diffusion of hydrophobic compounds through its LPS envelope [
36]. From the three examined oils, NO was the least effective (≈1.6-fold lower than the remainder oils). The EOs antimicrobial activity is attributed to the presence of several low molecular weight phenols, terpenes and aldoketones within their composition [
47]. Hence, the higher ZoI formed by TTO or CLO could be explained by the presence of volatile compounds. TTO encompasses terpinen-4-ol (40%), γ-terpinene (20%), α-terpinene (10%), 1,8-cineole (3%), α-pinene (3%) and limonene (1%) (
v/
v). TTO comprises in its formulation eugenol (80%), β-caryophyllene (4%), benzyl benzoate (4%), cinnamaldehyde (3%), linalool (2%) and α-terpinene (1%) (
v/
v). All these compounds are known to contribute significantly for the EOs antimicrobial activity [
26,
47,
48]. NO is known to possess terpinen-4-ol, γ-terpinene, α-terpinene, 1,8-cineole, α-pinene, limonene, β-caryophyllene and linalool, but with a major contribution of 1,8-cineole (60%) (
v/
v) to its antimicrobial activity. 1,8-cineole is known to exert lower antimicrobial action than terpinen-4-ol or eugenol [
28,
49,
50,
51,
52].
3.2. MICs
The obtained MICs of the selected antimicrobial agents for each bacterium are shown in
Table 2. MICs evaluation showed that the selected antimicrobial agents were active against all tested bacteria, which fairly agrees with the data obtained from the agar-well diffusion studies
From the entire list of tested agents, the MICs of the AgNPs were the highest (4000–1250 μg/mL). The main mechanism of action of AgNPs against bacteria requires the attachment and interaction of multiple NPs to the cell surface [
53]. This induces the disruption of the bacteria membrane functions and dissipation of the proton motive force. Due to their high surface-to-volume ratio, small AgNPs of few nanometers may even alter the morphology of the cell wall, increasing their diffusion towards the intracellular space, ultimately leading to the cell death [
54]. Here, even though PVP was used as a dispersant agent to produce AgNPs, there was still a large tendency to form agglomerates in colloidal dispersions (
Figure 1). Consequently, a large number of NPs were expected to be attracted and immobilized along the membrane of each bacterium to induce a bactericidal effect. Between the tested organisms,
P. aeruginosa was the most susceptible to the AgNPs action. It has been postulated that Gram-negative bacteria are more susceptible to AgNPs because AgNPs positive charges interact with the outer LPS membrane with more affinity than with the Gram-positive cell wall, which is thought to have fewer interaction sites [
55]. This effect, however, was not verified on the
E. coli, which MIC was equal to
S. aureus and
S. epidermidis.
As expected, vancomycin was more effective against Gram-positive than Gram-negative bacteria, with MICs being 120-fold lower [
2]. The pexiganan MICs were also consistent with the ZoI findings, being in a range close to that reported in the literature [
17,
18]. On the contrary, the LL37 action was found to be superior against Gram-negative bacteria, even though the ZoIs were more evident against the Gram-positive ones (
Table 1). Again, these results imply the difficulty of the peptide in diffusing through the solid media [
46]. Regarding the tested bacteria, the LL37 ability to act more effectively against the
E. coli and
P. aeruginosa bacteria is related to its electrostatic interaction and its structure. It has been reported that LL37s first interaction step is promoted by its electrostatic attraction to lipid A and to phosphate groups linked to sugar residues of LPS [
56]. Subsequently, α-helix peptides, such as LL37, generally act via a membranolytic mechanism. The helix formation allows an optimal spatial arrangement of the aliphatic side chains for membrane insertion. Strong hydrophobic interactions are formed between these chains and the lipid layer of Gram-negative bacteria, stabilizing the AMP helical conformation, thus reducing main-chain hydrophobicity and allowing for a deeper and easier insertion into the bilayer [
57,
58].
As shown in
Table 2, the EOs displayed variable levels of MICs for each tested microorganism. CLO had the lowest MICs (19.7–39.3 μg/mL) from the group, while NO had the highest (137.0–365.2 μg/mL). These findings corroborate the ZoI examinations (
Table 1). The EOs differences in chemical composition and presence of more effective low molecular weight antibacterial compounds on CLO than on NO, exerts a major role in their antibacterial activity efficacy [
47,
48,
49]. To the best of the authors’ knowledge, NO has not yet been tested against these specific strains. Still, in other cases, MICs have been reported around 300 and 500 μg/mL [
28]. Regarding the TTO MICs, even though they are slightly superior to those reported in the literature, the pattern of efficiency remains:
P. aeruginosa <
S. epidermidis <
S. aureus <
E. coli [
26].
3.3. Kill-Time Analysis
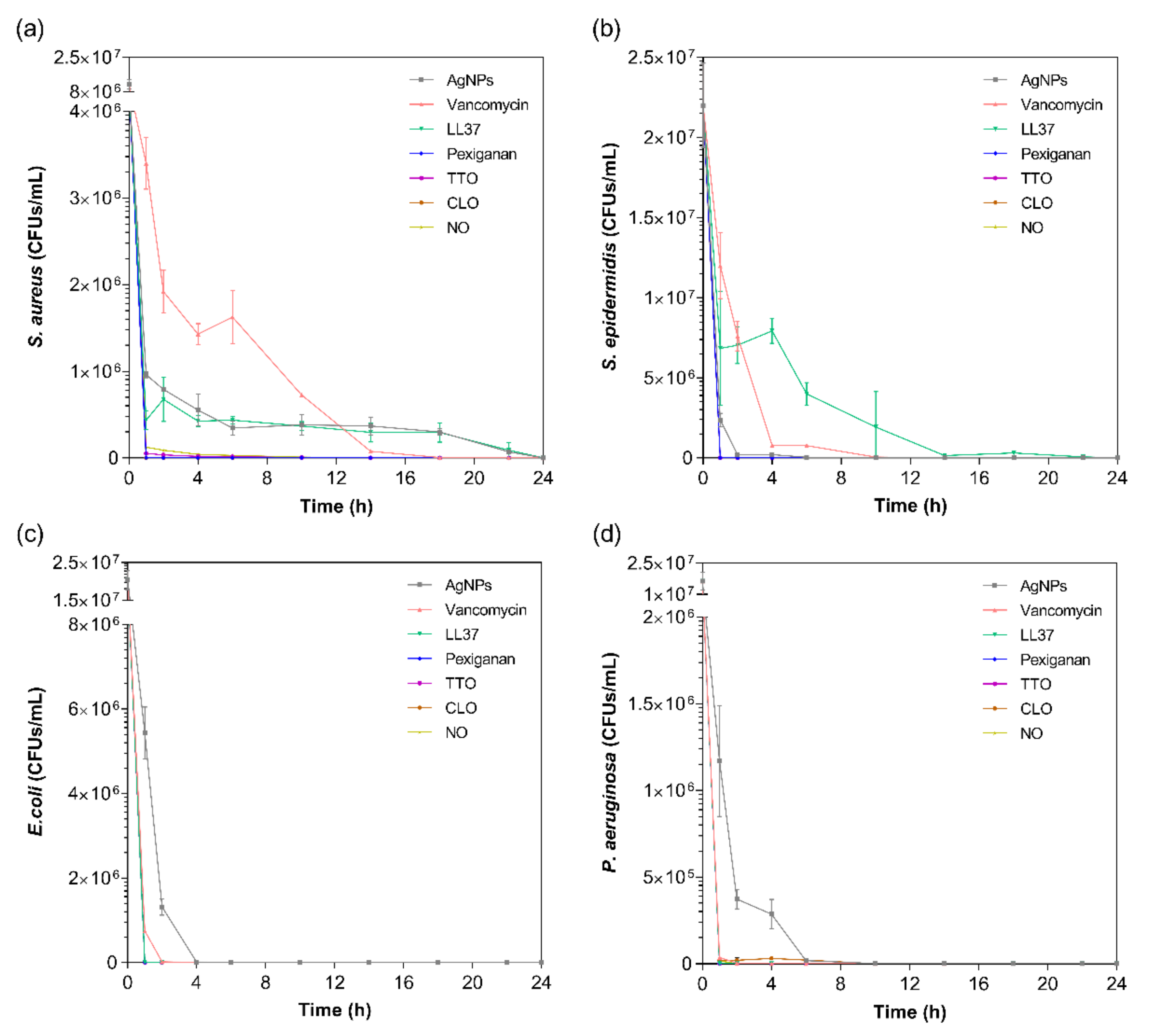
The kill-time kinetics for each agent was determined by the number of remaining viable cells at specific time points, namely 1, 2, 4, 6, 10, 14, 18, 22 and 24 h (
Figure 2). Although MIC values are commonly used to predict the antibacterial action of any agent, such data does not consider the exposure and action time of the agent against each bacterium. As such, the kill-time kinetics was used to unravel the antibacterial potency of the tested compounds over time.
For all bacteria/agent combinations, bactericidal action was observed from the first hour of contact. In fact, the action of the pexiganan, TTO, CLO and NO was so immediate that after 2 h of contact, very little bacteria remained (≈3 × 10
4 CFUs/mL of
S. aureus with TTO, ≈2 × 10
4 CFUs/mL of
P. aeruginosa with CLO, and ≈3 × 10
4 CFUs/mL of
S. aureus with NO; the remainder were all killed at this point). The main bactericidal action of the AMP pexiganan results from irreversible membrane-disruptive damage, which based on the mechanism of action of magainin (precursor), is expected to exert its antimicrobial action very quickly, within the first moments of interaction [
59]. Our data is consistent with this information and with previous reports [
17]. In fact, all bacteria were dead after 1 h of contact, with no regrowth being observed within the 24 h tested. Regarding the EOs, the susceptibility pattern for each bacterium did not appear to predict the activity of the EO. For instance, even though TTO required a very small concentration to kill
S. aureus, it took 6 h for this bacterium to be eliminated, whereas TTO only required 1 h to eradicate the other bacteria. The same occurred with NO. On its turn, CLO followed the pattern of MIC concentrations, requiring 6 h to kill the
P. aeruginosa and less than 1 h for the other bacteria. The EOs mechanism of action relies on their inherent hydrophobicity, which enables them to accumulate in the cell membrane, disturbing its structure and functionality, and causing an increase of permeability [
36,
60]. Despite sharing a similar membrane and cell wall structure and disposition, it is known that
E. coli and
P. aeruginosa possess distinct lipid and protein composition and concentration [
61]. This most likely is the main factor for the observed MIC differences between these bacteria. Even though it is not yet clear at which stage of bacteria development the EOs are the most effective, it is generally accepted that they stimulate cell autolysis in exponential and stationary cell phases [
62]. Our findings demonstrate that exponentially growing cells are very susceptible to the EOs’ action.
In case of the LL37, the action was quicker against Gram-negative bacteria compared to Gram-positive ones. It has been shown that permeabilization of the cytoplasmic membrane of Gram-positive bacteria by LL37 is considerably slower than against Gram-negative [
57,
58]. Interestingly, vancomycin also had a faster action against Gram-negative bacteria than against Gram-positive, even though all available data up until now has revealed its higher effectiveness towards the former. This may have happened due to the differences in MIC values. While
S. aureus’ and
S. epidermidis’ MIC was only 7.8 μg/mL, 1000 μg/mL of the antibiotic was necessary to kill
E. coli and
P. aeruginosa. Several studies have shown that the bactericidal action, namely kill-time kinetics, of a given antimicrobial agent is dependent on the concentration to which the microorganisms are exposed [
17,
63]. For the four bacteria, AgNPs was the agent that took the longest time to eliminate the entirety of CFUs. As seen in
Figure 1, AgNPs clusters prevented its homogenous dispersion within the solution, which implies that AgNPs were not evenly available to bind to sites at the bacteria membrane. As AgNPs are only capable of disrupting the bacteria cell membrane after proper binding, subsequently infiltrating the cytosol to induce cell death [
54], this limited distribution may have required additional time than the free-state, non-clustered molecules, characteristic of the other tested agents.
3.4. Cell-Wall Disruption: Mechanisms of Action
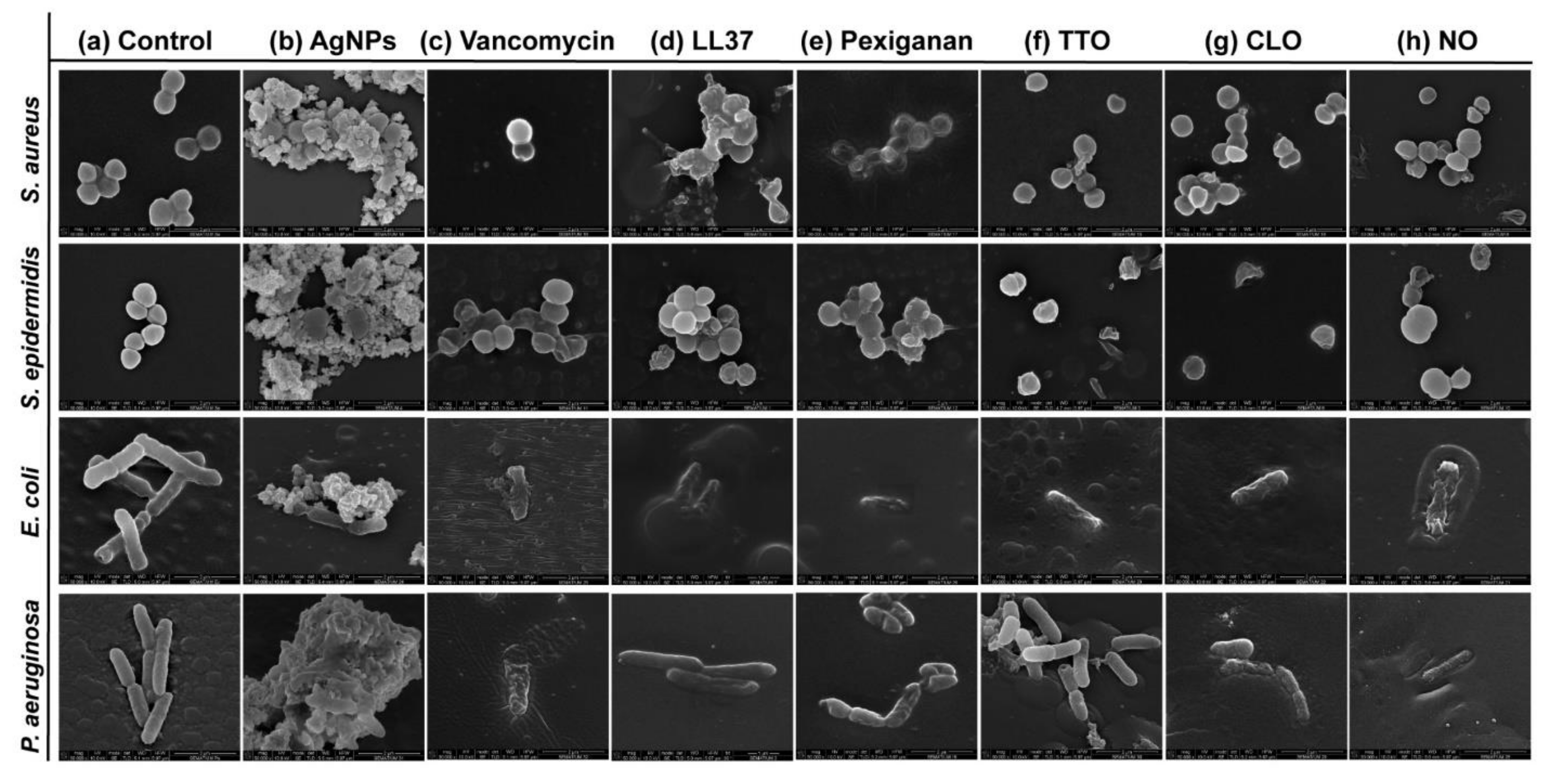
Possible mechanisms of membrane interaction and disruption of the tested bacteria have been observed via SEM imaging through exposition to the selected agents at half of the MICs concentrations, for 24 h.
Figure 3 shows the morphology of the bacteria with and without contact with AgNPs, vancomycin and selected AMPs and EOs. As expected, the control (without agent) of the
S. aureus and
S. epidermidis bacteria revealed a coccoid-shaped conformation with a smooth and uninterrupted surface. Both cell types tended to be arranged in grape-like clusters and cell propagation was recurrently observed [
64,
65]. The morphology of the
E. coli and
P. aeruginosa bacteria was also very similar, with both displaying a rod-shaped architecture.
E. coli cells are typically 2.0 μm long and 0.25–1.0 μm in diameter.
P. aeruginosa cells present similar dimensions and, in many cases, polar flagella, which endows the bacteria with motility, may also be evident [
66,
67,
68]. Here, however, that was not the case. Gram-negative control cells presented an even distribution, displaying multiple cells undergoing polar binary fission.
There are two mechanisms of action widely accepted for AgNPs, the contact killing and the ion-mediated killing. Contact killing is clearly evidenced in all tested microorganisms (
Figure 3b). The positively charged AgNPs are attracted to the negatively charged bacteria surface, enabling NP attachment along the cell surface. This action induces physical changes in the bacterial membrane, compromising its integrity and inducing the diffusion of NPs towards the intracellular space. Here, AgNPs species, such as Ag
+ ions, are released and interact effectively with specific functional groups in proteins, consequently inhibiting intracellular metabolic functions and causing protein denaturation. At this point, the cellular content will leak, ultimately leading to the cell death [
69,
70]. This effect is particularly evident against the
P. aeruginosa bacteria, as the rod-shaped morphology is barely evident in most cells and the cell content is already fused with the AgNPs clusters. Additionally, AgNPs are also capable of producing high levels of reactive oxygen species (ROS) and free radical species that may interact electrostatically with the cell wall, generating a charge superior to its tensile strength, therefore also compromising its integrity [
71].
Direct inhibition of the Atl amidase domain (major domain in staphylococcus bacteria cell wall), due to vancomycin-induced inhibition of cell wall synthesis, causes defects in the cell morphology and alters cell membrane permeability (
S. aureus in
Figure 3c), ultimately leading to autolysin-triggered cell rupture, release of cell content and death (
S. epidermidis in
Figure 3c) [
40,
41,
42]. Even though the mode of action against
S. aureus and
S. epidermidis is similar,
Figure 3c recorded the alterations induced by vancomycin at two stages, an earlier for
S. aureus and a more advanced for
S. epidermidis. This occurs because vancomycin requires more time to kill the first bacteria than the second (
Figure 2), thus allowing for the extrusion and reduction of cell content to occur more intensively in the
S. epidermidis upon 24 h exposure. The same explanation can be applied to the Gram-negative bacteria. The large size of this glycopeptide precludes it from being capable of penetrating the outer membrane of Gram-negative bacteria and inducing morphology changes and autolysin-triggered cell rupture, as happens in Gram-positive bacteria [
43,
44,
45]. We postulate that because of the superior concentration of vancomycin (1000 μg/mL) required to kill these bacteria and its fast action, these molecules accumulate along the surface of the bacterium, isolating it from the media and respective nutrients, and providing enough steric hindrance to prevent peptidoglycan synthesis. Hence, starving the bacteria may trigger a set of events somewhat similar to those characteristics of the Gram-positive bacteria that culminate in cell rupture, release of cell content and death. As the kill-time kinetics is so fast, after 24 h exposure, it was only possible to capture fragments of individual cell membranes and residues of cell content for
E. coli and a very advanced deformed morphology for
P. aeruginosa.
AMPs are unique biomolecules which mode of action may be divided into direct killing (membrane and non-membrane target) and immune modulation. LL37-treated
S. aureus displayed clear irregular protruding structures, to an extent that at least some bacteria appeared to have extruded cytoplasm and become embedded by exudate. In the case of
S. epidermidis, morphological changes were easily distinguished, with a small leakage of cytoplasm content also being perceived within the bacteria cluster. LL37 performs its bactericidal action by electrostatic binding of its cationic molecules to the outer surface of the bacterial cell. This peptide uses the carpet-like mechanism, in which the AMP coat the microbial membrane up to saturation, after which point wormholes are formed, or the toroidal mechanism of action, in which after binding to the phospholipid head groups, the AMP aligns and inserts into the membrane and cluster into unstructured bundles that span the membrane and generate channels from each of the intracellular content leaks [
12,
14]. LL37 is amphiphilic in nature with hydrophobic and hydrophilic residues aligned on opposite sides of the peptide. This facilitates their penetration through the cell membrane [
72], which results in inhibition of nucleic acid and protein biosynthesis, followed by leakage of the cell cytoplasm into the extracellular space, causing bacteria death [
73]. Although the action of LL37 against Gram-negative bacteria is very similar to that described against Gram-positive bacteria, the rate at which cytoplasmic permeabilization occurs is superior. The peptide α-helix structure forms strong hydrophobic interactions with the outer membrane and its LPS and O-antigen layers of the Gram-negative bacteria, which quickly saturates, thus allowing for a deeper and faster insertion into the bilayer [
57,
58]. The halting of growth occurs shortly after the translocation of LL37 across the outer membrane into the periplasmic space, and is followed by the rapid interference with the synthesis of the nascent curved cell envelope (the outer membrane, cytoplasmic membrane, peptidoglycan layer and LPS layer) and its intracellular organelles [
74]. These phenomena may explain the more advanced state of deformation/decomposition registered by the
E. coli and
P. aeruginosa bacteria after 24 h exposure to the LL37 (
Figure 3d). Here, a substantial decrease in signal intensity correlates to cells being depleted of intracellular organelles, in a bed of organic matter (very easily identified for
E. coli). Just as LL37, pexiganan also exert its antibacterial effect by forming toroidal pores in the bacterial membrane [
75]. The cationic AMP with divalent cation binding sites disrupts the arrangement of the hydrophobic and hydrophilic sections of the bilayer, inducing a local curvature which alters the morphological appearance of the cell (evident in all bacteria from
Figure 3e). As the pores are transient upon disintegration, pexiganan can translocate to the inner cytoplasmic leaflet entering the cytoplasm and potentially targeting intracellular components [
17,
76]. As the antimicrobial action of AMPs, including pexiganan, is related to its availability, bacteria exposed to a higher concentration of pexiganan were more prone to disintegrate and release their cellular content during the 24 h contact. This was particularly clear on the
E. coli, for which MIC was the highest (62.5 μg/mL). As observed, AMP kills bacteria very quickly, within the first 2 h of exposure (
Figure 2), by physically disrupting the cell membrane, which is a highly conserved structure, thus the development of resistance may not be an immediate concern, which potentiates further research into its clinical application [
77,
78]. In fact, all mutagenesis attempts to induce pexiganan resistance in
E. coli and
S. aureus failed [
79].
It is generally accepted that EOs act primarily against the cell cytoplasmic membrane of the microorganism. Their inherent hydrophobicity is an important characteristic that enables them to accumulate within the cell membrane, disturbing its structure and functionality, and causing an increase of permeability. Leakage of intracellular components and impairment of microbial functions can then occur, ultimately causing cell death [
36,
60]. Even though in
S. aureus the action of TTO appeared to have only compromised the cell wall with little cell content being released, in
S. epidermidis, its effect is very pronounced with the complete disintegration of the cell membrane and substantial leakage to the extracellular space. Here, cell lysis is clear. This difference in behavior between staphylococcus bacteria may be correlated with the kill-time kinetics of the EO. While
S. aureus withstands viable cells for 6 h, the other tested microorganisms were all eliminated within 1 h. In fact, disintegration of the cell wall, leakage of intracellular components and cell lysis are especially noticeable in
E. coli and
P. aeruginosa (
Figure 3f). Another explanation relies on the TTO action mode against
S. aureus. Data suggests that the primary mechanism of action against this bacterium may not be just gross cell wall damage, as it happens with the other bacteria, but rather a combination of the weakening of the cell wall and subsequent rupture of the cytoplasmic membrane due to osmotic pressure with the impairment of microbial autolytic enzyme systems, which eventually induce a delayed death [
50,
51]. Obvious detrimental effects on the cell membrane morphology were also shown when bacteria were treated with CLO and NO. Microstructural observations demonstrated these EOs’ capacity to increase cell permeability, distorting the cell membrane integrity and generating holes or wrinkles. The latter were particularly clear on the Gram-negative bacteria, possibly due to their outer cell membrane and thin peptidoglycan wall. An incomplete and deformed shape was observed in some
S. aureus and
S. epidermidis cells. Cell shrinkage and blebbing-like architectures were also detected among these microorganisms. Interestingly, intracellular leakage was only observed on
S. aureus, even though rupture and lysis of membranes with a “breaking-in-half”-like deformation was predominant in
S. epidermidis. CLO is composed of 80% eugenol, and its antibacterial action can be attributed to a double bond in the α,β positions of the side chain and a methyl group located in the γ position. Typically, eugenol exhibits higher activity against Gram-negative bacteria than Gram-positive bacteria [
50]. Deformation of the bacterial cell wall of Gram-negative bacteria is evident upon exposure to CLO. It appears that the EO surrounds the cells, isolating them, for an effective permeabilization of the membrane. Indeed, since these bacteria are relatively more resistant to hydrophobic biomolecules, to overcome their impermeability, EOs rely on the organisms isolation to slowly traverse through the outer wall porins [
50,
80]. This phenomenon is also evident on the Gram-negative bacteria treated with NO. Still, the differences in concentration between CLO and NO necessary to accomplish such a task (
Table 2) may be accompanied by a different mechanism of action against these bacteria. As the concentration of phenolic compounds in NO is smaller than on CLO [
47,
48,
49], the first may rely on the interference with enzymes involved in the production of energy to induce cell lysis, while the second may denature proteins present at the intracellular space.

 ,
, 







{kind=link}
{kind=link}
{kind=link}



























