Wingless/Wnt Signaling in Intestinal Development, Homeostasis, Regeneration and Tumorigenesis: A Drosophila Perspective

{kind=link}
{kind=link}
Abstract
:1. The Canonical Wnt/β-Catenin Signaling Pathway
1.1. Wnt/β-Catenin Signaling Pathway
1.2. Wnt/β-Catenin Signaling in Development and Disease
2. Wnt/β-Catenin Signaling in Mammalian Intestinal Physiology and Pathology
3. The Drosophila Adult Gut: A Powerful Model for Studying Wnt Signaling
4. Wg Signaling in the Drosophila Gut: Development, Homeostasis, Regeneration, and Tumorigenesis
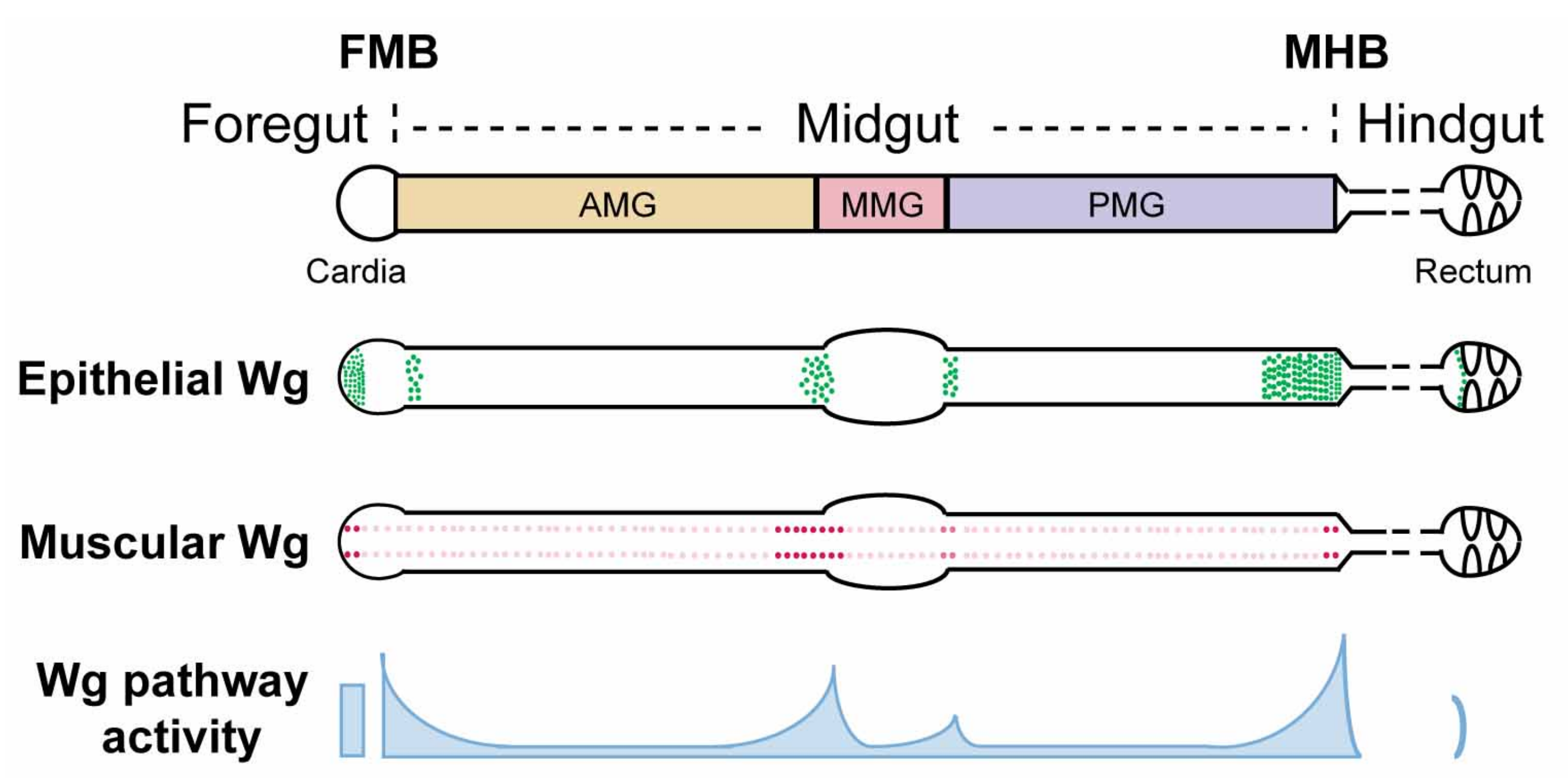
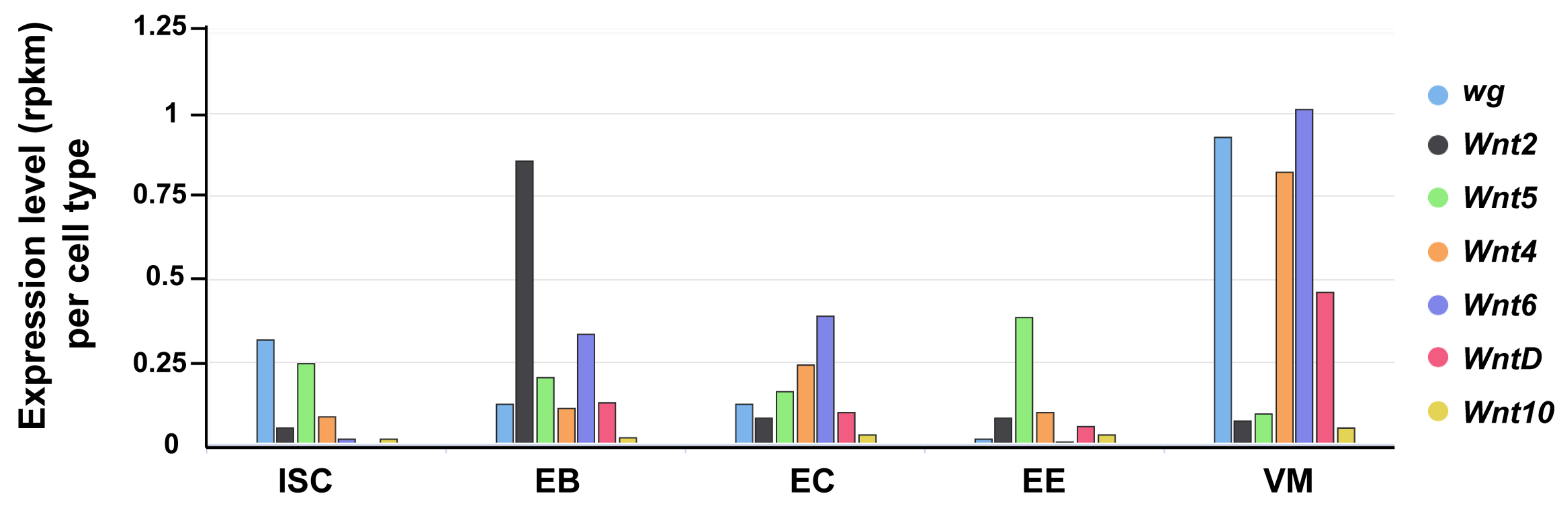
4.1. Wg Is Expressed at Major Compartmental Boundaries in the Adult Midgut
4.2. Graded Activation of Wg Signaling at Major Compartment Boundaries in the Drosophila Midgut
4.3. Wg Directs Pattern Formation during Drosophila Gut Development
4.3.1. Wg Signaling in Formation of the Adult Intestinal Midgut/Hindgut Boundary during Pupation
4.3.2. Wg Signaling in Formation of the Foregut/Midgut Boundary of the Adult Gut during Development
4.3.3. Wg Signaling in Embryonic and Larval Gut Development
4.4. Wg Signaling Regulates ISC Self-Renewal/Maintenance and Proliferation in the Drosophila Adult Gut during Homeostasis
4.5. Wg Signaling in Adult Midgut and Hindgut Regeneration Following Injury
4.6. Hyperactivation of Wg Signaling Due to Loss of Apc: Initiation and Progression of Tumorigenesis in the Drosophila Gut
4.6.1. Initiation of Intestinal Tumorigenesis upon Loss of Apc
4.6.2. Progression of Intestinal Tumorigenesis Following Apc Loss
5. The Drosophila Gut as a Powerful In Vivo Context to Test Novel Therapeutic Agents and Novel Wnt Pathway Components
5.1. At the Receptor Level: The Signalosome
5.2. In the Cytoplasm: Tankyrase
5.3. In the Nucleus: Earthbound and Erect Wing
6. Crosstalk between Wg Signaling and Other Signaling Pathways in the Drosophila Gut
7. Conclusions
8. Future Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gammons, M.; Bienz, M. Multiprotein complexes governing wnt signal transduction. Curr. Opin. Cell Biol. 2017, 51, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R.; Clevers, H. Wnt/beta-catenin signaling, disease, and emerging therapeutic modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Saito-Diaz, K.; Chen, T.W.; Wang, X.; Thorne, C.A.; Wallace, H.A.; Page-McCaw, A.; Lee, E. The way wnt works: Components and mechanism. Growth Factors 2013, 31, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Stamos, J.L.; Weis, W.I. The beta-catenin destruction complex. Cold Spring Harbor Perspect. Biol. 2013, 5, a007898. [Google Scholar] [CrossRef] [PubMed]
- Langton, P.F.; Kakugawa, S.; Vincent, J.P. Making, exporting, and modulating wnts. Trends Cell Biol. 2016, 26, 756–765. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Cong, F. Novel regulation of wnt signaling at the proximal membrane level. Trends Biochem. Sci. 2016, 41, 773–783. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, B.T.; He, X. Frizzled and lrp5/6 receptors for wnt/beta-catenin signaling. Cold Spring Harbor Perspect. Biol. 2012, 4, a007880. [Google Scholar] [CrossRef] [PubMed]
- De Bruine, Z.J.; Xu, H.E.; Melcher, K. Assembly and architecture of the wnt/beta-catenin signalosome at the membrane. Br. J. Pharmacol. 2017, 174, 4564–4574. [Google Scholar] [CrossRef] [PubMed]
- Li, V.S.; Ng, S.S.; Boersema, P.J.; Low, T.Y.; Karthaus, W.R.; Gerlach, J.P.; Mohammed, S.; Heck, A.J.; Maurice, M.M.; Mahmoudi, T.; et al. Wnt signaling through inhibition of beta-catenin degradation in an intact axin1 complex. Cell 2012, 149, 1245–1256. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, J.P.; Emmink, B.L.; Nojima, H.; Kranenburg, O.; Maurice, M.M. Wnt signalling induces accumulation of phosphorylated beta-catenin in two distinct cytosolic complexes. Open Biol. 2014, 4, 140120. [Google Scholar] [CrossRef] [PubMed]
- Masuda, T.; Ishitani, T. Context-dependent regulation of the beta-catenin transcriptional complex supports diverse functions of wnt/beta-catenin signaling. J. Biochem. 2017, 161, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, A.B.; Sinha, A.; Fan, V.B.; Cadigan, K.M. The wnt transcriptional switch: Tle removal or inactivation? BioEssays News Rev. Mol. Cell. Dev. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Cadigan, K.M.; Waterman, M.L. Tcf/lefs and wnt signaling in the nucleus. Cold Spring Harbor Perspect. Biol. 2012, 4, a007906. [Google Scholar] [CrossRef]
- Loh, K.M.; van Amerongen, R.; Nusse, R. Generating cellular diversity and spatial form: Wnt signaling and the evolution of multicellular animals. Dev. Cell 2016, 38, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Holstein, T.W. The evolution of the wnt pathway. Cold Spring Harbor Perspect. Biol. 2012, 4, a007922. [Google Scholar] [CrossRef] [PubMed]
- Petersen, C.P.; Reddien, P.W. Wnt signaling and the polarity of the primary body axis. Cell 2009, 139, 1056–1068. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.; Loh, K.M.; Nusse, R. Stem cell signaling. An integral program for tissue renewal and regeneration: Wnt signaling and stem cell control. Science 2014, 346, 1248012. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, K.; Clevers, H. Wnt/beta-catenin signaling in adult mammalian epithelial stem cells. Dev. Biol. 2017, 428, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Lien, W.H.; Fuchs, E. Wnt some lose some: Transcriptional governance of stem cells by wnt/beta-catenin signaling. Genes Dev. 2014, 28, 1517–1532. [Google Scholar] [CrossRef] [PubMed]
- Bastakoty, D.; Young, P.P. Wnt/beta-catenin pathway in tissue injury: Roles in pathology and therapeutic opportunities for regeneration. FASEB J. 2016, 30, 3271–3284. [Google Scholar] [CrossRef] [PubMed]
- Majidinia, M.; Aghazadeh, J.; Jahanban-Esfahlani, R.; Yousefi, B. The roles of wnt/beta-catenin pathway in tissue development and regenerative medicine. J. Cell. Physiol. 2017. [Google Scholar] [CrossRef]
- Clevers, H.; Nusse, R. Wnt/beta-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt signaling in cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef] [PubMed]
- Baron, R.; Kneissel, M. Wnt signaling in bone homeostasis and disease: From human mutations to treatments. Nat. Med. 2013, 19, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Krausova, M.; Korinek, V. Wnt signaling in adult intestinal stem cells and cancer. Cell. Signal. 2014, 26, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Schepers, A.; Clevers, H. Wnt signaling, stem cells, and cancer of the gastrointestinal tract. Cold Spring Harbor Perspect. Biol. 2012, 4, a007989. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in globocan 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Potten, C.S.; Morris, R.J. Epithelial stem-cells invivo. J. Cell Sci. 1988, 10, 45–62. [Google Scholar] [CrossRef]
- Kosinski, C.; Li, V.S.; Chan, A.S.; Zhang, J.; Ho, C.; Tsui, W.Y.; Chan, T.L.; Mifflin, R.C.; Powell, D.W.; Yuen, S.T.; et al. Gene expression patterns of human colon tops and basal crypts and bmp antagonists as intestinal stem cell niche factors. Proc. Natl. Acad. Sci. USA 2007, 104, 15418–15423. [Google Scholar] [CrossRef] [PubMed]
- van de Wetering, M.; Sancho, E.; Verweij, C.; de Lau, W.; Oving, I.; Hurlstone, A.; van der Horn, K.; Batlle, E.; Coudreuse, D.; Haramis, A.P.; et al. The beta-catenin/tcf-4 complex imposes a crypt progenitor phenotype on colorectal cancer cells. Cell 2002, 111, 241–250. [Google Scholar] [CrossRef]
- Gregorieff, A.; Pinto, D.; Begthel, H.; Destree, O.; Kielman, M.; Clevers, H. Expression pattern of wnt signaling components in the adult intestine. Gastroenterology 2005, 129, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Farin, H.F.; Jordens, I.; Mosa, M.H.; Basak, O.; Korving, J.; Tauriello, D.V.; de Punder, K.; Angers, S.; Peters, P.J.; Maurice, M.M.; et al. Visualization of a short-range wnt gradient in the intestinal stem-cell niche. Nature 2016, 530, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Fevr, T.; Robine, S.; Louvard, D.; Huelsken, J. Wnt/beta-catenin is essential for intestinal homeostasis and maintenance of intestinal stem cells. Mol. Cell. Biol. 2007, 27, 7551–7559. [Google Scholar] [CrossRef] [PubMed]
- Ireland, H.; Kemp, R.; Houghton, C.; Howard, L.; Clarke, A.R.; Sansom, O.J.; Winton, D.J. Inducible cre-mediated control of gene expression in the murine gastrointestinal tract: Effect of loss of beta-catenin. Gastroenterology 2004, 126, 1236–1246. [Google Scholar] [CrossRef] [PubMed]
- Kuhnert, F.; Davis, C.R.; Wang, H.T.; Chu, P.; Lee, M.; Yuan, J.; Nusse, R.; Kuo, C.J. Essential requirement for wnt signaling in proliferation of adult small intestine and colon revealed by adenoviral expression of dickkopf-1. Proc. Natl. Acad. Sci. USA 2004, 101, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.; Gregorieff, A.; Begthel, H.; Clevers, H. Canonical wnt signals are essential for homeostasis of the intestinal epithelium. Genes Dev. 2003, 17, 1709–1713. [Google Scholar] [CrossRef] [PubMed]
- Korinek, V.; Barker, N.; Moerer, P.; van Donselaar, E.; Huls, G.; Peters, P.J.; Clevers, H. Depletion of epithelial stem-cell compartments in the small intestine of mice lacking tcf-4. Nat. Genet. 1998, 19, 379–383. [Google Scholar] [CrossRef] [PubMed]
- van Es, J.H.; Haegebarth, A.; Kujala, P.; Itzkovitz, S.; Koo, B.K.; Boj, S.F.; Korving, J.; van den Born, M.; van Oudenaarden, A.; Robine, S.; et al. A critical role for the wnt effector tcf4 in adult intestinal homeostatic self-renewal. Mol. Cell. Biol. 2012, 32, 1918–1927. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.A.; Kakitani, M.; Zhao, J.; Oshima, T.; Tang, T.; Binnerts, M.; Liu, Y.; Boyle, B.; Park, E.; Emtage, P.; et al. Mitogenic influence of human r-spondin1 on the intestinal epithelium. Science 2005, 309, 1256–1259. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Vries, R.G.; Snippert, H.J.; van de Wetering, M.; Barker, N.; Stange, D.E.; van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef] [PubMed]
- The Cancer Genome Atlas Network. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337. [Google Scholar]
- Groden, J.; Thliveris, A.; Samowitz, W.; Carlson, M.; Gelbert, L.; Albertsen, H.; Joslyn, G.; Stevens, J.; Spirio, L.; Robertson, M.; et al. Identification and characterization of the familial adenomatous polyposis-coli gene. Cell 1991, 66, 589–600. [Google Scholar] [CrossRef]
- Kinzler, K.W.; Nilbert, M.C.; Su, L.K.; Vogelstein, B.; Bryan, T.M.; Levy, D.B.; Smith, K.J.; Preisinger, A.C.; Hedge, P.; McKechnie, D.; et al. Identification of fap locus genes from chromosome 5q21. Science 1991, 253, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Nishisho, I.; Nakamura, Y.; Miyoshi, Y.; Miki, Y.; Ando, H.; Horii, A.; Koyama, K.; Utsunomiya, J.; Baba, S.; Hedge, P. Mutations of chromosome 5q21 genes in fap and colorectal cancer patients. Science 1991, 253, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Wood, L.D.; Parsons, D.W.; Jones, S.; Lin, J.; Sjoblom, T.; Leary, R.J.; Shen, D.; Boca, S.M.; Barber, T.; Ptak, J.; et al. The genomic landscapes of human breast and colorectal cancers. Science 2007, 318, 1108–1113. [Google Scholar] [CrossRef] [PubMed]
- Bass, A.J.; Lawrence, M.S.; Brace, L.E.; Ramos, A.H.; Drier, Y.; Cibulskis, K.; Sougnez, C.; Voet, D.; Saksena, G.; Sivachenko, A.; et al. Genomic sequencing of colorectal adenocarcinomas identifies a recurrent vti1a-tcf7l2 fusion. Nat. Genet. 2011, 43, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Dong, X.; Mai, M.; Seelan, R.S.; Taniguchi, K.; Krishnadath, K.K.; Halling, K.C.; Cunningham, J.M.; Boardman, L.A.; Qian, C.; et al. Mutations in axin2 cause colorectal cancer with defective mismatch repair by activating beta-catenin/tcf signalling. Nat. Genet. 2000, 26, 146–147. [Google Scholar] [CrossRef] [PubMed]
- Morin, P.J.; Sparks, A.B.; Korinek, V.; Barker, N.; Clevers, H.; Vogelstein, B.; Kinzler, K.W. Activation of beta-catenin-tcf signaling in colon cancer by mutations in beta-catenin or apc. Science 1997, 275, 1787–1790. [Google Scholar] [CrossRef] [PubMed]
- Seshagiri, S.; Stawiski, E.W.; Durinck, S.; Modrusan, Z.; Storm, E.E.; Conboy, C.B.; Chaudhuri, S.; Guan, Y.; Janakiraman, V.; Jaiswal, B.S.; et al. Recurrent r-spondin fusions in colon cancer. Nature 2012, 488, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, S.M.; Fearon, E.R. Axin1 and axin2 variants in gastrointestinal cancers. Cancer Lett. 2014, 355, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Giannakis, M.; Hodis, E.; Jasmine Mu, X.; Yamauchi, M.; Rosenbluh, J.; Cibulskis, K.; Saksena, G.; Lawrence, M.S.; Qian, Z.R.; Nishihara, R.; et al. Rnf43 is frequently mutated in colorectal and endometrial cancers. Nat. Genet. 2014, 46, 1264–1266. [Google Scholar] [CrossRef] [PubMed]
- Lum, L.; Clevers, H. Cell biology. The unusual case of porcupine. Science 2012, 337, 922–923. [Google Scholar] [CrossRef] [PubMed]
- Koo, B.K.; Spit, M.; Jordens, I.; Low, T.Y.; Stange, D.E.; van de Wetering, M.; van Es, J.H.; Mohammed, S.; Heck, A.J.; Maurice, M.M.; et al. Tumour suppressor rnf43 is a stem-cell e3 ligase that induces endocytosis of wnt receptors. Nature 2012, 488, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.X.; ** reveals regulatory complexity in the adult drosophila midgut. Cell Rep. 2015, 12, 346–358. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, B.; Miguel-Aliaga, I. The digestive tract of drosophila melanogaster. Annu. Rev. Genet. 2013, 47, 377–404. [Google Scholar] [CrossRef] [PubMed]
- Buchon, N.; Osman, D. All for one and one for all: Regionalization of the drosophila intestine. Insect Biochem. Mol. 2015, 67, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Micchelli, C.A.; Perrimon, N. Evidence that stem cells reside in the adult drosophila midgut epithelium. Nature 2006, 439, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Ohlstein, B.; Spradling, A. The adult drosophila posterior midgut is maintained by pluripotent stem cells. Nature 2006, 439, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Biteau, B.; Jasper, H. Slit/robo signaling regulates cell fate decisions in the intestinal stem cell lineage of drosophila. Cell Rep. 2014, 7, 1867–1875. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Hou, S.X. Enteroendocrine cells are generated from stem cells through a distinct progenitor in the adult drosophila posterior midgut. Development 2015, 142, 644–653. [Google Scholar] [CrossRef] [PubMed]
- Zielke, N.; Korzelius, J.; van Straaten, M.; Bender, K.; Schuhknecht, G.F.; Dutta, D.; ** drosophila midgut stem cells. Dev. Biol. 2016, 416, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Takashima, S.; Younossi-Hartenstein, A.; Ortiz, P.A.; Hartenstein, V. A novel tissue in an established model system: The drosophila pupal midgut. Dev. Genes Evol. 2011, 221, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Ohlstein, B. Stem cell regulation. Bidirectional notch signaling regulates drosophila intestinal stem cell multipotency. Science 2015, 350. [Google Scholar] [CrossRef] [PubMed]
- Aghajanian, P.; Takashima, S.; Paul, M.; Younossi-Hartenstein, A.; Hartenstein, V. Metamorphosis of the drosophila visceral musculature and its role in intestinal morphogenesis and stem cell formation. Dev. Biol. 2016, 420, 43–59. [Google Scholar] [CrossRef] [PubMed]
- Klapper, R. The longitudinal visceral musculature of drosophila melanogaster persists through metamorphosis. Mech. Dev. 2000, 95, 47–54. [Google Scholar] [CrossRef]
- Kuroda, J.; Nakamura, M.; Yoshida, M.; Yamamoto, H.; Maeda, T.; Taniguchi, K.; Nakazawa, N.; Hatori, R.; Ishio, A.; Ozaki, A.; et al. Canonical wnt signaling in the visceral muscle is required for left-right asymmetric development of the drosophila midgut. Mech. Dev. 2012, 128, 625–639. [Google Scholar] [CrossRef] [PubMed]
- Takashima, S.; Murakami, R. Regulation of pattern formation in the drosophila hindgut by wg, hh, dpp, and en. Mech. Dev. 2001, 101, 79–90. [Google Scholar] [CrossRef]
- Hoppler, S.; Bienz, M. Two different thresholds of wingless signalling with distinct developmental consequences in the drosophila midgut. EMBO J. 1995, 14, 5016–5026. [Google Scholar] [PubMed]
- Lee, W.C.; Beebe, K.; Sudmeier, L.; Micchelli, C.A. Adenomatous polyposis coli regulates drosophila intestinal stem cell proliferation. Development 2009, 136, 2255–2264. [Google Scholar] [CrossRef] [PubMed]
- Strand, M.; Micchelli, C.A. Quiescent gastric stem cells maintain the adult drosophila stomach. Proc. Natl. Acad. Sci. USA 2011, 108, 17696–17701. [Google Scholar] [CrossRef] [PubMed]
- Cordero, J.B.; Stefanatos, R.K.; Myant, K.; Vidal, M.; Sansom, O.J. Non-autonomous crosstalk between the jak/stat and egfr pathways mediates apc1-driven intestinal stem cell hyperplasia in the drosophila adult midgut. Development 2012, 139, 4524–4535. [Google Scholar] [CrossRef] [PubMed]
- Buchon, N.; Broderick, N.A.; Lemaitre, B. Gut homeostasis in a microbial world: Insights from drosophila melanogaster. Nat. Rev. Microbiol. 2013, 11, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Tian, A.; Jiang, J. Intestinal stem cell response to injury: Lessons from drosophila. Cell. Mol. Life Sci. CMLS 2016, 73, 3337–3349. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hodgson, J.J.; Buchon, N. Drosophila as a model for homeostatic, antibacterial, and antiviral mechanisms in the gut. PLoS Pathog. 2017, 13, e1006277. [Google Scholar] [CrossRef] [PubMed]
- Losick, V.P.; Fox, D.T.; Spradling, A.C. Polyploidization and cell fusion contribute to wound healing in the adult drosophila epithelium. Curr. Biol. CB 2013, 23, 2224–2232. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, Y.; Hayashi, S.; Levine, A.; Wieschaus, E. Regulation of armadillo by a drosophila apc inhibits neuronal apoptosis during retinal development. Cell 1998, 93, 1171–1182. [Google Scholar] [CrossRef]
- Ahmed, Y.; Nouri, A.; Wieschaus, E. Drosophila apc1 and apc2 regulate wingless transduction throughout development. Development 2002, 129, 1751–1762. [Google Scholar] [PubMed]
- Akong, K.; Grevengoed, E.E.; Price, M.H.; McCartney, B.M.; Hayden, M.A.; DeNofrio, J.C.; Peifer, M. Drosophila apc2 and apc1 play overlap** roles in wingless signaling in the embryo and imaginal discs. Dev. Biol. 2002, 250, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Rubinfeld, B.; Souza, B.; Polakis, P.; Wieschaus, E.; Levine, A.J. A drosophila homolog of the tumor suppressor gene adenomatous polyposis coli down-regulates beta-catenin but its zygotic expression is not essential for the regulation of armadillo. Proc. Natl. Acad. Sci. USA 1997, 94, 242–247. [Google Scholar] [CrossRef] [PubMed]
- McCartney, B.M.; Dierick, H.A.; Kirkpatrick, C.; Moline, M.M.; Baas, A.; Peifer, M.; Bejsovec, A. Drosophila apc2 is a cytoskeletally-associated protein that regulates wingless signaling in the embryonic epidermis. J. Cell Biol. 1999, 146, 1303–1318. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Waltzer, L.; Bienz, M. A new drosophila apc homologue associated with adhesive zones of epithelial cells. Nat. Cell Biol. 1999, 1, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Cordero, J.; Vidal, M.; Sansom, O. Apc as a master regulator of intestinal homeostasis and transformation: From flies to vertebrates. Cell Cycle 2009, 8, 2926–2931. [Google Scholar] [CrossRef] [PubMed]
- Suijkerbuijk, S.J.; Kolahgar, G.; Kucinski, I.; Piddini, E. Cell competition drives the growth of intestinal adenomas in drosophila. Curr. Biol. CB 2016, 26, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhao, R.; Huang, P.; Yang, F.; Quan, Z.; Xu, N.; ** of genes predisposing to idiopathic generalized epilepsy. Hum. Mol. Genet. 1995, 4, 1201–1207. [Google Scholar] [CrossRef] [PubMed]
- DeSimone, S.M.; White, K. The drosophila erect wing gene, which is important for both neuronal and muscle development, encodes a protein which is similar to the sea urchin p3a2 DNA binding protein. Mol. Cell. Biol. 1993, 13, 3641–3649. [Google Scholar] [CrossRef] [PubMed]
- Fazio, I.K.; Bolger, T.A.; Gill, G. Conserved regions of the drosophila erect wing protein contribute both positively and negatively to transcriptional activity. J. Biol. Chem. 2001, 276, 18710–18716. [Google Scholar] [CrossRef] [PubMed]
- Virbasius, C.A.; Virbasius, J.V.; Scarpulla, R.C. Nrf-1, an activator involved in nuclear-mitochondrial interactions, utilizes a new DNA-binding domain conserved in a family of developmental regulators. Genes Dev. 1993, 7, 2431–2445. [Google Scholar] [CrossRef] [PubMed]
- Major, M.B.; Roberts, B.S.; Berndt, J.D.; Marine, S.; Anastas, J.; Chung, N.; Ferrer, M.; Yi, X.; Stoick-Cooper, C.L.; von Haller, P.D.; et al. New regulators of wnt/beta-catenin signaling revealed by integrative molecular screening. Sci. Signal. 2008, 1, ra12. [Google Scholar] [CrossRef] [PubMed]
- Pangon, L.; Ng, I.; Giry-Laterriere, M.; Currey, N.; Morgan, A.; Benthani, F.; Tran, P.N.; Al-Sohaily, S.; Segelov, E.; Parker, B.L.; et al. Jrk is a positive regulator of beta-catenin transcriptional activity commonly overexpressed in colon, breast and ovarian cancer. Oncogene 2016, 35, 2834–2841. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Lucchetta, E.; Rafel, N.; Ohlstein, B. Maintenance of the adult drosophila intestine: All roads lead to homeostasis. Curr. Opin. Genet. Dev. 2016, 40, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Biteau, B.; Hochmuth, C.E.; Jasper, H. Jnk activity in somatic stem cells causes loss of tissue homeostasis in the aging drosophila gut. Cell Stem Cell 2008, 3, 442–455. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, A.; Benchabane, H.; Ahmed, Y. Wingless/Wnt Signaling in Intestinal Development, Homeostasis, Regeneration and Tumorigenesis: A Drosophila Perspective. J. Dev. Biol. 2018, 6, 8. https://doi.org/10.3390/jdb6020008
Tian A, Benchabane H, Ahmed Y. Wingless/Wnt Signaling in Intestinal Development, Homeostasis, Regeneration and Tumorigenesis: A Drosophila Perspective. Journal of Developmental Biology. 2018; 6(2):8. https://doi.org/10.3390/jdb6020008
Chicago/Turabian StyleTian, Ai, Hassina Benchabane, and Yashi Ahmed. 2018. "Wingless/Wnt Signaling in Intestinal Development, Homeostasis, Regeneration and Tumorigenesis: A Drosophila Perspective" Journal of Developmental Biology 6, no. 2: 8. https://doi.org/10.3390/jdb6020008