Plants initiate several mechanisms such as stimulation of antioxidant system, accumulation of osmolytes or secondary metabolites, heat shock proteins to resist the harmful effect of heat stress through maintenance of cellular homeostasis and repair of damaged membranes and proteins. These mechanisms are associated with production of various hormones and signalling molecules that help in adapting to heat-induced adversities. Kaya et al. [
46] reported that interactive effect of H
2S and NO resulted in reducing oxidative stress and cadmium uptake by enhancing the antioxidative enzyme system and mineral nutrients uptake. At low concentration, H
2S affects plant development and growth by playing a major role in abiotic and biotic stress responses [
27,
62]. Hydrogen sulfide has gained much attention because it is a vital mode of sulfur metabolism in organisms and has increased special significance owed to its signalling properties, besides the essentiality of S requirement is known under both abiotic and biotic stress [
63]. Under conditions of stress when ROS accumulation causes oxidative stress, the increased H
2S helps in reducing the ROS via enhancing the non-enzymatic and enzymatic ways. In our study also H
2S evolution increased under heat stress which might have signalled for increase in antioxidative mechanism because reduction of H
2S by hypotaurin decreased antioxidative enzymes, photosynthesis, and growthunder heat stress. The rescue mechanism of H
2S also involves signal that regulates stomatal movement, increases GSH, redox ratio, along with GSH-associated genes expression under chilling stress [
64]. H
2S relates through H
2O
2 and was found to upregulate ascorbate-glutathione cycle which acted as the downstream signal for H
2S regulation on ROS [
65,
66]. Exogenous H
2S application on plant seedlings was reported to inhibit oxidative stress by reducing the MDA activity, electrolyte leakage and proline [
66,
67] and protected photosynthesis and stablized the chloroplast structure [
68]. Under heat stress accumulation of H
2S has been reported which could be associated with the acquisition of stress tolerance in plants [
69,
70]. Besides, exogenously applied H
2S can induce cross-adaptation to multiple stresses, indicating its potential as a signal molecule in cross-adaptation in plants [
70,
71].
The interaction of H
2S with various other signalling molecules has also been reported. Hydrogen sulfide with Ca
2+ and CaM effectively alleviated heat-induced damage to plants via increasing the H
2S accumulation [
72] and H
2S-induced heat tolerance required Ca
2+ transport toward cytoplasm then intracellular CaM coordinates it [
73]. Methylglyoxal (MG) which resembles H
2S contributes in abiotic stress response and application of MG and/or NaHS enhanced maize seedlings response to heat stress and both showed positive interaction [
74]. A crosstalk of H
2S signals with carbon monoxide, salicylic acid (SA), abscisic acid (ABA), and ethylene has also been reported [
75,
76,
77,
78], and these are found to prompt H
2S-producing enzymes activation and endogenous H
2S accretion under high-temperature stress. Hydrogen sulfide interacts with ethylene under osmotic stress [
79], with ABA for closure of stomata [
78], with NO for heat tolerance [
4] with SA for lead tolerance [
47], with H
2O
2 and brassinosteroid (BR) for stomatal closure [
80] and with melatonin for salt and iron deficiency tolerance [
46].
While undergoing various studies we found that melatonin interaction with H
2S under heat stress has not been worked out, although antioxidants and photosynthesis are shown to be influenced with H
2S application. We still do not know how H
2S affects carbohydrate metabolism under heat stress and its relationship with melatonin. Melatonin is known as a biopromoter as it regulates numerous physiological methods and improves heat stress resistance in plants [
81,
82]. It is a small molecular weight indoleamine hormone explicitly being measured as a candidate phytohormone due to its various responses under biotic and abiotic stresses. Its level increases upon plant’s exposure to abiotic stresses and it scavenges ROS molecules and enhances activity of antioxidant enzyme, content of metabolite, photosynthetic efficiency and regulates stress transcription features and other metabolites and signalling molecules [
83]. However, the information on how melatonin regulates carbohydrate metabolism and photosynthesis in heat stress in wheat and its interplay with H
2S in this mechanism is scanty. In the present study, melatonin enhanced plants’ photosynthetic potential by increasing carbohydrate accumulation with enhanced antioxidative enzymes activity. The influence of melatonin was found to be dependent on H
2S evolution as evidenced that the reduction of H
2S reducedphotosynthesis and growth in wheat.
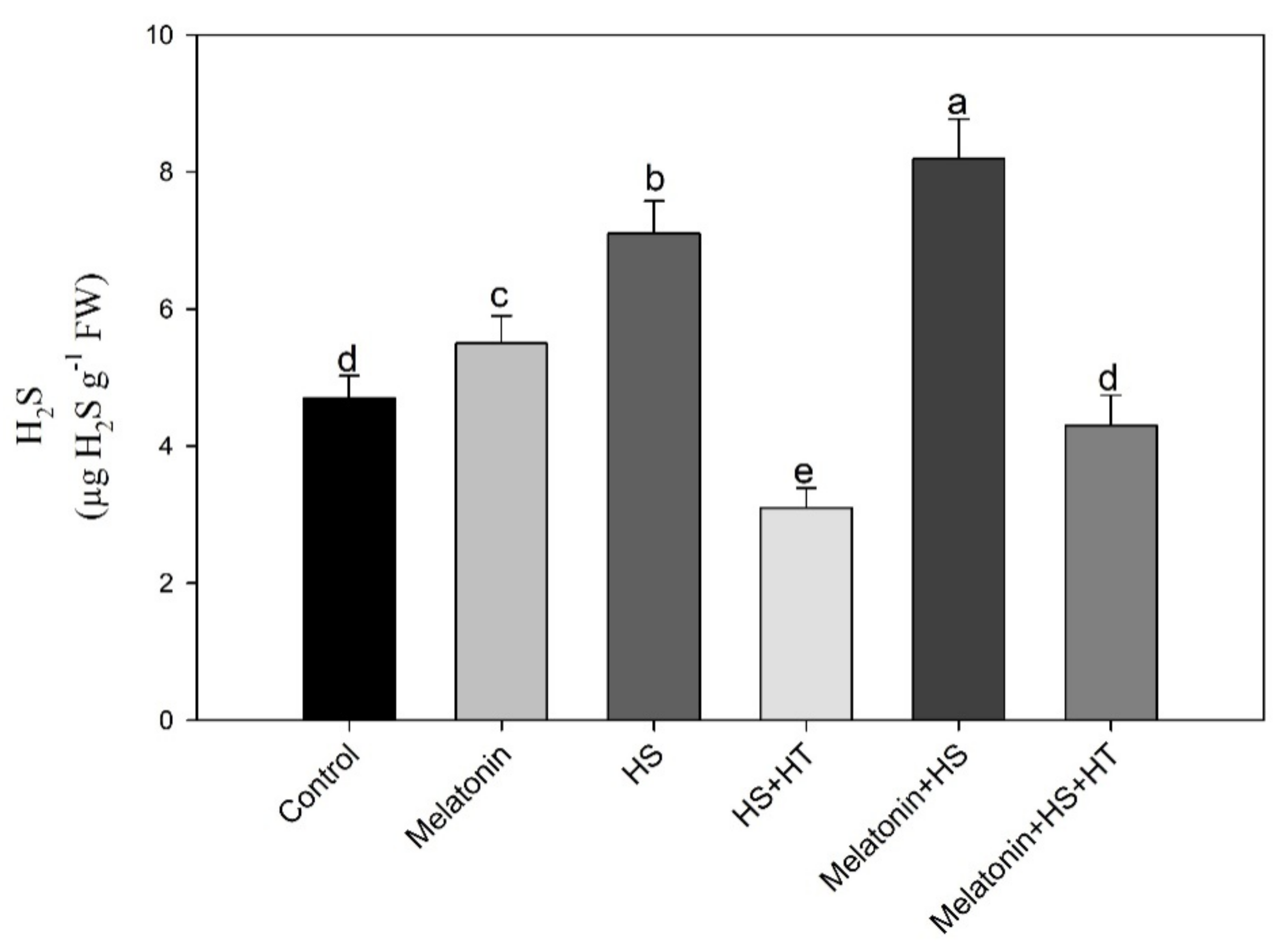
Similar responses in reduction of all studied photosynthetic traits were observed when H2S was reduced with hypotaurine under heat stress showing H2S signalling under heat stress as an adaptation strategy against heat adversities on plants.
4.1. Melatonin Increases H2S Evolution in Wheat under Heat Stress
Exogenous application of melatonin further increased H
2S evolution which was induced under heat stress. Previous studies have reported that heat stress induces H
2S evolution [
4]. The interrelationship between melatonin and H
2S has been stated in some researches. Kaya et al. [
46] found that melatonin imparted resistance to join deficiency of iron and salt-stress which was intervened through both NO and H
2S. These molecules went about as downstream signal molecule in melatonin actuated resilience. Exogenous melatonin treatment under salt stress modulated the endogenous H
2S level and absolutely upregulated the L-cysteine desulfhydrase activity under salt stress in tomato seedlings [
25]. Turk and Erdal [
84] studied that melatonin application enhanced mineral element content in cold-stressed plants. Among studied nutrients, sulfur level also increased by melatonin under stress which acts as a building block of proteins, enzymes and vitamins. Sulfide is an important intermediate in sulfur metabolism and the amount of H
2S released has been correlated with sulfate supply to the plants [
71]. When hypotaurinewas supplemented to heat stressed plants with or without melatonin, reduction in H
2S level was observed and more reduction in plants under heat stress and without melatonin. With lower level of H
2S detected in heat stressed and hypotaurine treated plants, we observed greater reduction in all the parameters in heat stress and hypotaurine treatment.
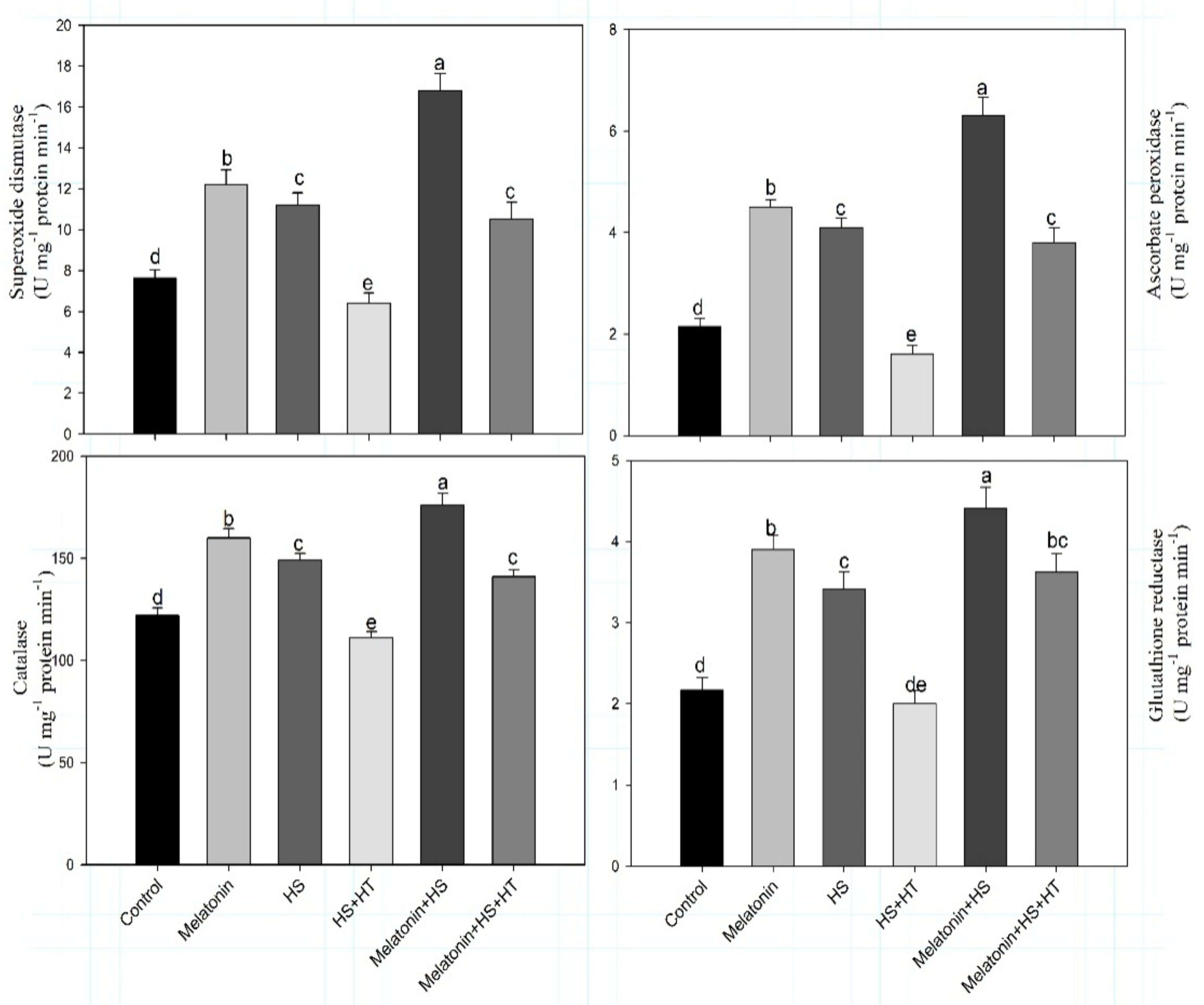
4.2. Melatonin Decreases Heat Stress-Induced Oxidative Stress by Enhancing the Antioxidative Machinery: The Effect Mediated by H2S
In this research, heat stress was found to increase oxidative stress supported by increased TBARS and H
2O
2 content which was reduced by the supplementation of melatonin. Melatonin supplementation increased the activity of antioxidative enzymes (CAT, SOD, APR, GR) which scavenges the excess generated ROS to reduce oxidative stress. Enhanced activity of antioxidative enzymes detoxifies excess ROS and reduces oxidative stress. Catalase activity increased under heat stress and more increase occurred with melatonin treatment under heat stress. Catalase and peroxidises play important role in regulating the level of intracellular H
2O
2 and convert it into H
2O regenerating NADP
+ that helps plant under stress conditions [
85]. Increase in CAT activity was found under heat stress in wheat; and heat tolerance in wheat genotypes was directly linked with the percent enhancement in antioxidantive enzymes including CAT, SOD and guaiacol peroxidase [
86]. Reports suggest that melatonin mediates several enzymes expression that detoxify excess H
2O
2 such as, CAT, glutathione/ascorbate reductases, peroxidases in addition to peroxiredoxins [
17,
87] and balances the ROS/RNS level through antioxidant action. Either endogenous or exogenous supplied melatonin induces its own synthesis presenting melatonin as a specific regulator, an anti-stress manager and a plant master regulator [
88,
89]. Siddiqui et al. [
90] reported that melatonin induces photosynthetic enzymes, antioxidative system, proline metabolism and carbohydrate level under salt stress. Since supplementation of melatonin increases H
2S therefore, it was thought that melatonin action was via H
2S. Kaya et al. [
46] have studied crosstalk among H
2S and NO in melatonin-induced salt stress tolerance in pepper. NaHS pretreatment enhances cellular viability and was found to decrease MDA accumulation and electrolyte leakage [
73]. H
2S diminishes the harmful effect of oxidative stress under heat stress [
75]. In our study, we found that SOD, CAT, APX, GR activity amplified in stress and more increase was received with melatonin, but 47.6% reduction of H
2S activity with hytotaurine in melatonin and heat stress treatment reduced the increase in antioxidative enzymes that was observed in melatonin and heat-stressed plants. Similarly, Iqbal et al. [
4] reported that H
2S enhances antioxidative enzymes activity and ascorbate-glutathione cycle to scavenge excess ROS and reduce oxidative stress in wheat under heat stress. Although not much has been discussed on the interaction between melatonin and H
2S under heat studies, various studies support for a regulatory interaction either directly or indirectly. The relationship of melatonin, H
2S and NO was considered in regulation of fruit ripening [
91] and under salt stress by Kaya et al. [
46]. However, the present reported study is the first onepertaining to melatonin and H
2S under heat stress in wheat where melatonin increased the H
2S generation to scavenge excess ROS by increasing the antioxidative enzymes activity.
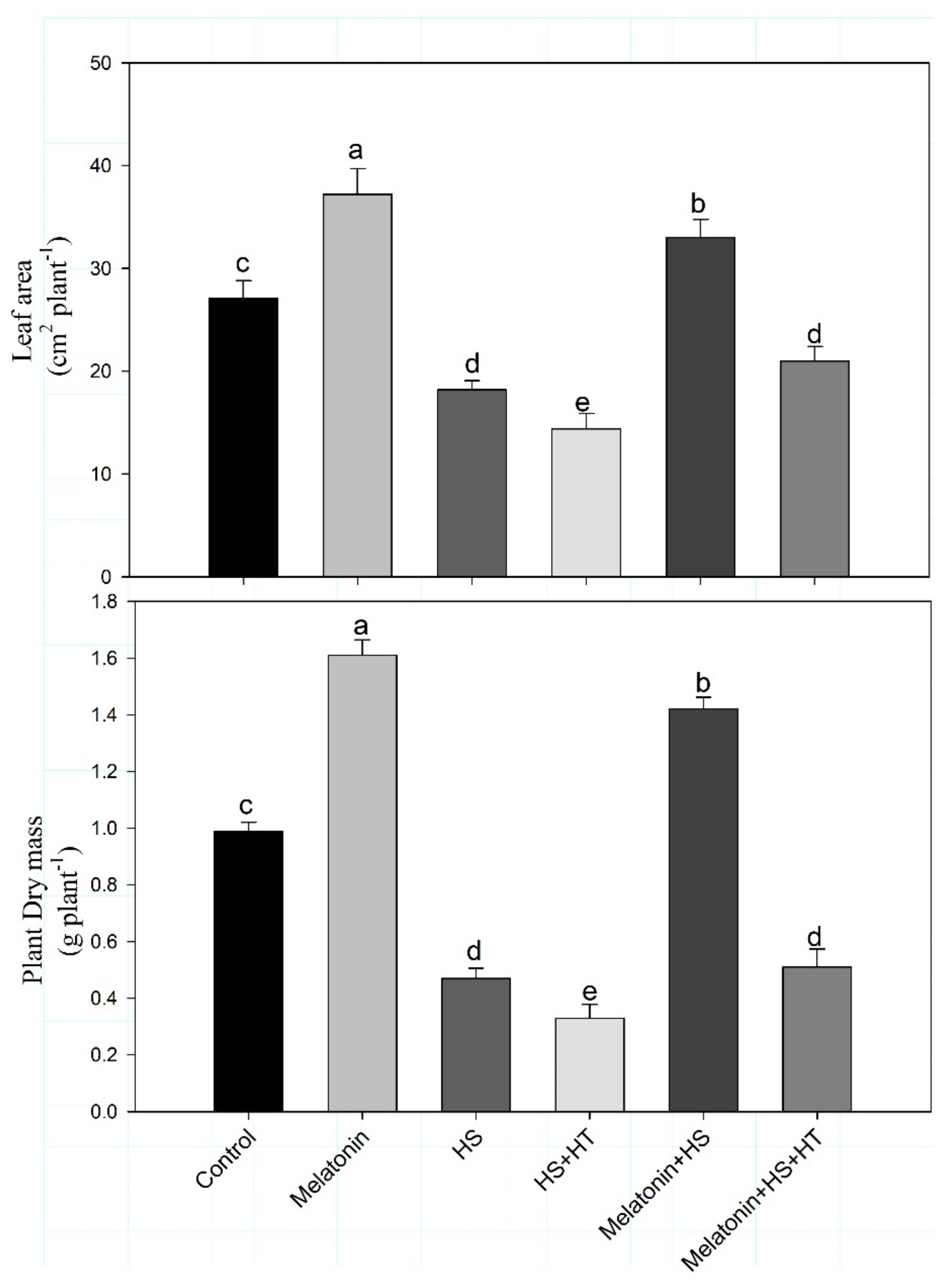
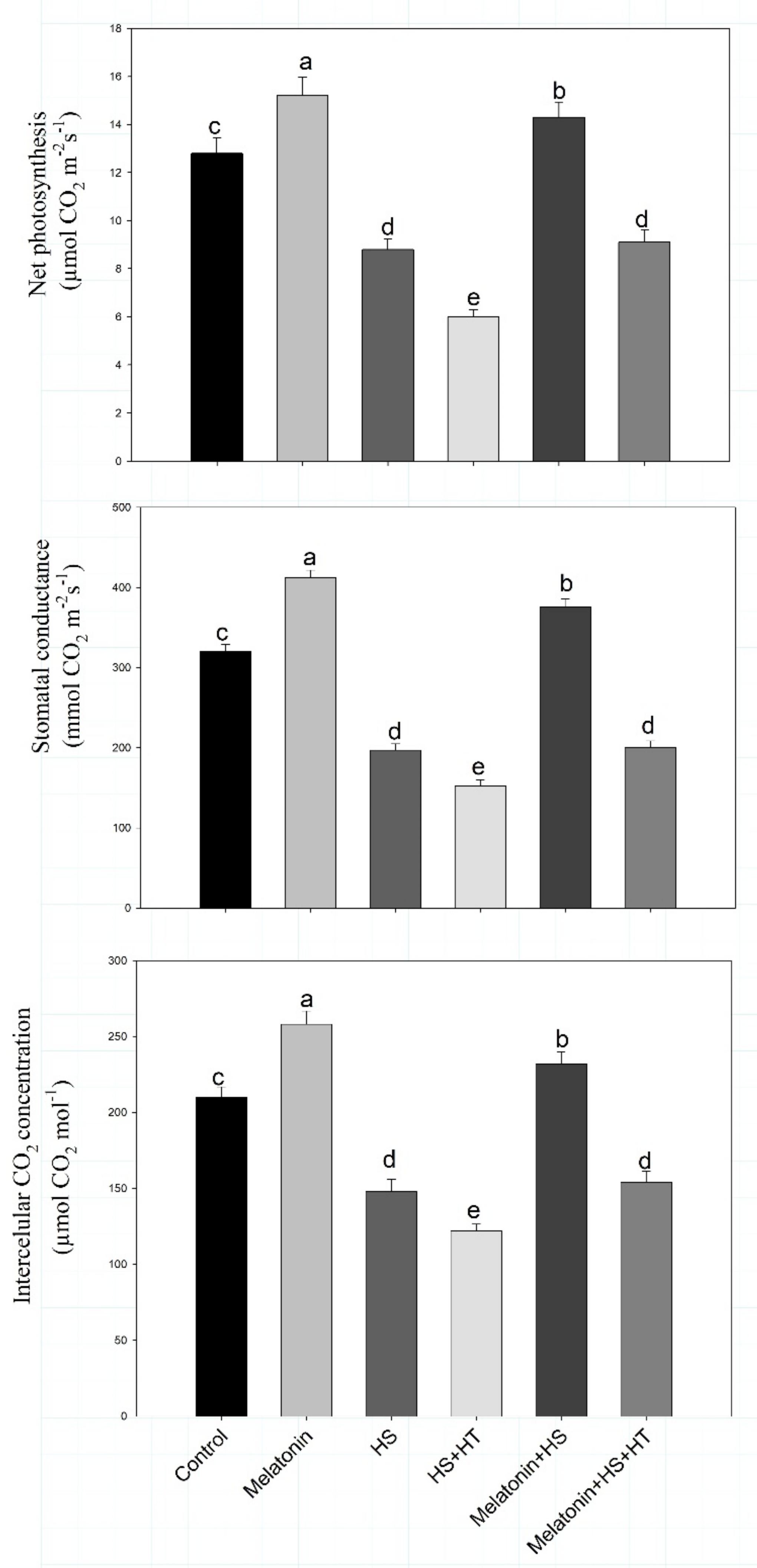
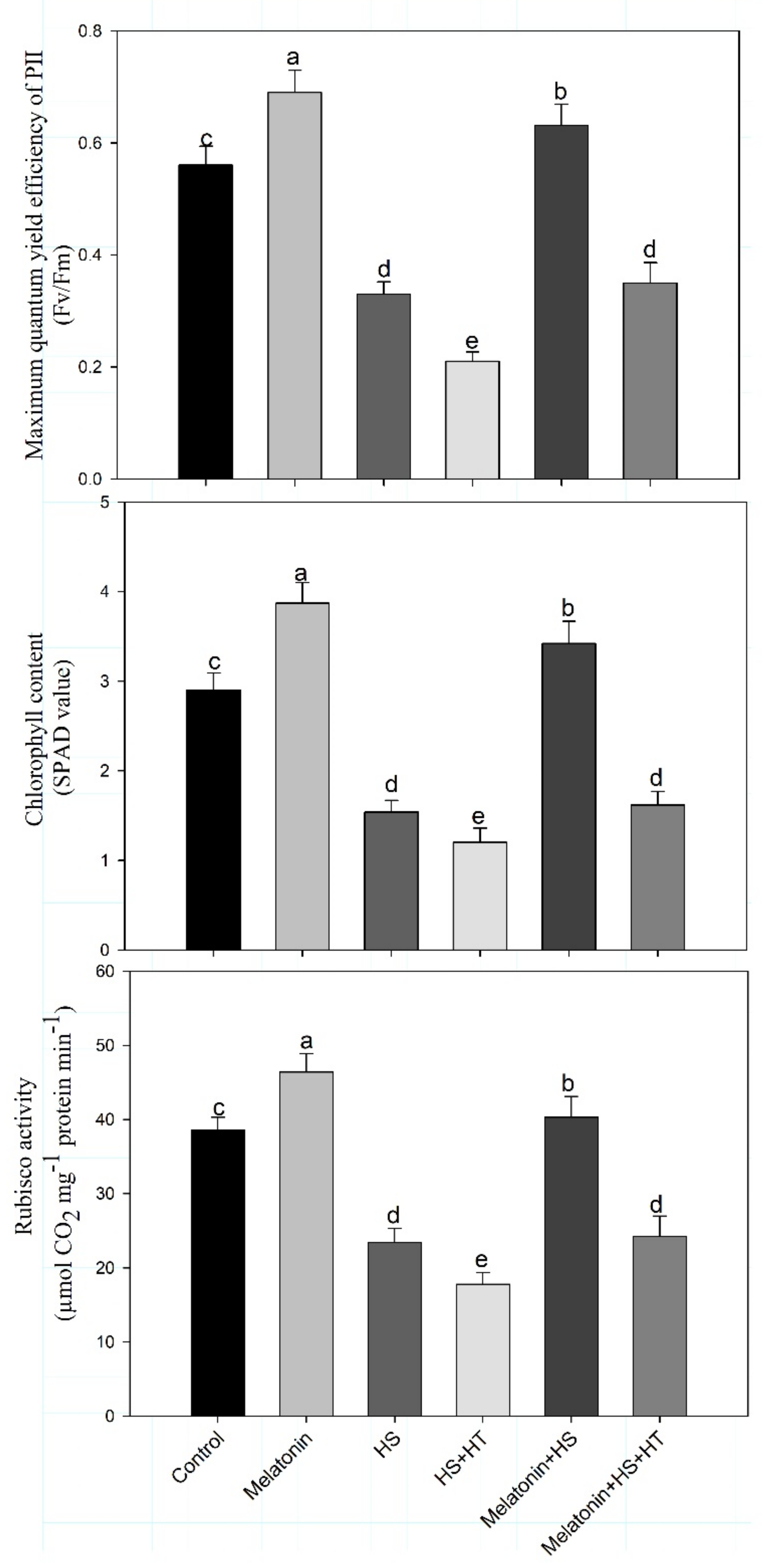
4.3. Impact of Melatonin on Photosynthesis and Carbohydrate Metabolism under Heat Stress: Reversal of the Effect by H2S Scavenger
Heat stress affects the process of photosynthesis [
4,
92] by altering the activities of enzymes in metabolism of carbon and it also affects sucrose synthesis and starch accumulation by modulating explicit genes of the carbohydrate metabolism pathway [
93]. The reduction in photosynthetic efficiency could be attributed to reduced activity of the Calvin cycle enzymes comprising Rubisco [
94]. We found decreased Rubisco activity under heat stress which was restored by melatonin application. Reduction in chlorophyll content also leads to reduced photosynthesis probably due to increased chlorophyllase activity [
95]. In the present study, the impact of melatonin was observed in reducing heat-induced photosynthetic reduction. Heat-stress reduced the content of chlorophyll, Rubisco activity in addition to efficiency of PSII and reduced stomatal conductance, intercellular CO
2 concentration and net photosynthesis. However, melatonin supply increased Rubisco, chlorophyll content and photosynthetic traits which were reduced by application of H
2S scavenger suggesting the involvement of H
2S in melatonin-induced increase in photosynthesis under heat stress. Individually both melatonin and H
2S are reported to enhance photosynthesis but here we found melatonin effect to be mediated by H
2S. Melatonin supplementation was found to increase photosynthesis efficiency by elevating Rubisco and fructose bisphosphatase enzyme activities under heat stress in tomato seedlings [
96].
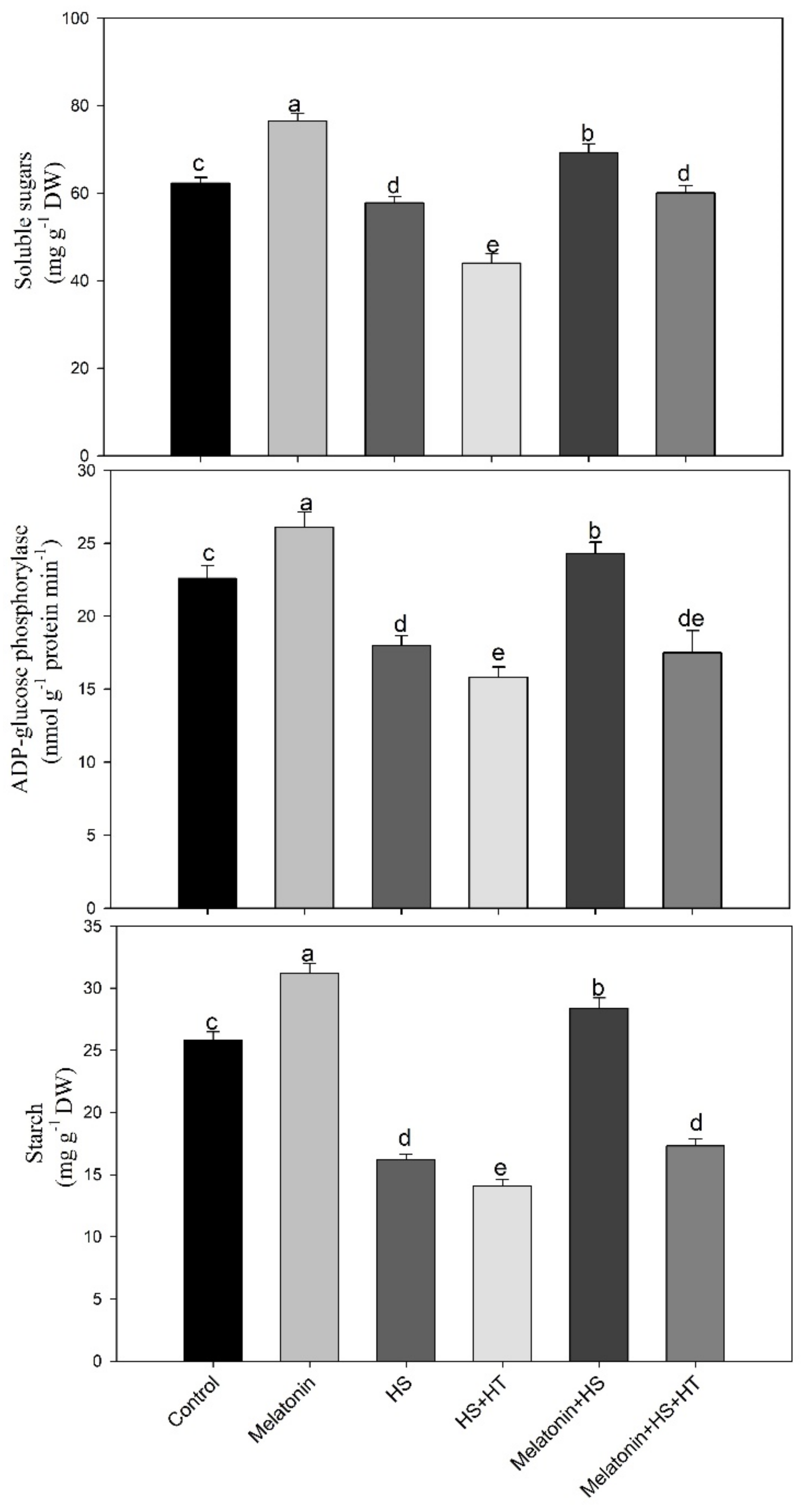
We observed that heat stress led to a decrease in accumulation of starch content and total soluble sugar. Exposure of plants to heat stress can be harmful to the starch synthesis enzymes activity and starch content [
97]. In several studies, ADP-Glucose pyrophosphorylase activity in maize, wheat, barley and rice were repressed under heat stress which reduced starch biosynthesis [
97,
98]. The present work reveals that the activity of ADP-Glucose pyrophosphorylase, one of the core rate limiting starch biosynthesis enzymes decreased in heat-stressed plants. Conversely, study also suggests that heat stress primes to rise in content of carbohydrates that helps to uphold cell turgor, helps in cell layers’ stabilization and prevents deterioration of protein [
99]. Melatonin supplementation in sugar starved suspension cells of
Nicotiana tabacum L. line Bright Yellow played regulatory role in carbohydrate metabolism byincreasing starch content through diverting the cell metabolism on gluconeogenesis whichled to carbohydrate synthesis from nonsugar precursors, like amino acids [
100]. In response to stress melatonin improves plants photosynthetic ability [
101,
102].
Soluble sugar acts as a signalling molecule that controls growth and metabolic processes and defence linked genes to regulate plant growth [
103,
104]. Thus, enhanced sugar accumulation is required for potential tolerance of plants and we observed rise in sucrose under heat stress which was reduced by melatonin supplementation when the stress was relieved and photosynthesis and starch increased. The degradation of starch under heat stress was supported by reduced ADP-Glucose phosphorylase activity then increased sucrose phosphate synthase and sucrose synthase activity which helped in utilization of sugars for plant growth. Melatonin led to enhancement of starch and sucrose content to alleviate heat induced oxidative stress by maintaining an osmotic balance and providing energy and food for growth. These actions of melatonin were blocked by H
2S scavenger which supported the role of H
2S in increasing carbohydrate accumulation under heat stress.
Heat tolerance requires the accumulation of sugars in plants and availability of carbohydrate (glucose and sucrose), is a significant physiological attribute for the resistance to heat [
105]. They reported decreased carbohydrate content under heat stress that was more in heat-susceptible genotype compared to tolerant one. In our study, the content of total soluble sugar decreased in heat stress, but by supplementation of melatonin under heat stress sugar accumulation increased.
Figure 9 highlights the important changes that occur in the carbohydrate metabolism under heat stress. It may be said that plants under heat stress increase their sucrose content by favouring the degradation of starch. The conversion of sucrose to starch is reduced with reduced activity of ADP-Glucose phosphorylase but increased sucrose phosphate activity increases sucrose content. We observed that the activity of invertase and sucrose synthase reduced which resulted in reduced total soluble sugars, also with reduced starch content. The interconversion of starch-sugar in leaf is necessary for the tolerance in plants to abiotic stress [
106].
Enhanced sucrose phosphate synthase activity under temperature stress helps in shifting the carbon grade starch to sucrose synthesis [
107]. Melatonin increased the activity of sucrose synthase and invertase under heat stress thus leading to increased mobilization of carbon source to demanding sink. Increased expression of sucrose phosphate synthase genes is reported to increase heat tolerance. Melatonin was found to effect sucrose-phosphate synthase and sucrose synthase by either protein activation or gene transcriptions and increased invertase activity by decreasing invertase inhibitor gene expression and thus affected growth [
108]. Lafta and Lorenzen [
109] reported that at high temperatures, improvement in the sucrose phosphate synthase genes expression reasons upsurge in the sucrose synthesis that improves tolerance of plants under slight heat stress. Alike result was seen in our study by increased sucrose and sucrose phosphate synthase activity in heat stress. Sucrose phosphate synthase activity further increased with melatonin treatment under heat stress, however, sucrose content decreased on melatonin treatment compared to heat-stressed plants although they were greater than control. The reason for this could be the utilization of sucrose with melatonin for plants growth and development with the release of stress. Khan et al. [
106] reported increased sucrose content to be aligned with decreased starch content under salt stress in tomato seedling roots. They found the effect of K
+ in salt stress alleviation was mediated by H
2S through its effect on carbohydrate metabolism and antioxidative defense system. The melatonin-induced increase in carbohydrate was reduced when hypotaurine was supplemented with melatonin suggesting that this carbohydrate accumulation by melatonin require H
2S for its action under heat stress.
The increase in invertase activity with melatonin helps in heat tolerance and this increase was mediated through H
2S. Siddiqui et al. [
90] stated that melatonin alleviated salt stress through osmoregulation via increasingthe content of total soluble carbohydrate and proline and upregulated carbohydrate metabolism. Increased invertase activity enhanced sucrose transport and was found to improve heat tolerance [
73].
Hydrogen sulfide was found to increase glucose utilization under heat stress in wheat and decreased heat-induced reduction in photosynthesis [
4]. NaHS treatment to
S. oleracea leaves increased accumulation of glucose and sucrose though it was lower than control; nevertheless, trehalose and fructose contents considerably increased above the control suggesting the role of trehalose and fructose in drought tolerance. Increased photosynthesis and PSII efficiency in
S. oleracea under drought stress on H
2S supplementation was due to modulation of sugar metabolism [
69]. Ye et al. [
74] found that H
2S increases photosynthesis by promoting Rubisco activity plus photosynthetic electron transport increases chlorophyll biosynthesis. It also modulates Fv/Fm ratio, chlorophyll content together with net photosynthetic rate, stomatal conductance, and transpiration rate [
65,
110]. Khan et al. [
106] reported that potassium and endogenous H
2S regulated sugar metabolism for initiating adaptive responses under salinity stress. Hydrogen sulfide was found to alleviate salt stress and increased sucrose synthase, sucrose phosphate synthase, invertase activity and decreased starch content.
Although not much study has been done on the role of H
2S in regulating carbohydrate, here we observed that the reduction of H
2S by 47.6% by its scavenger resulted in decreased melatonin function on carbohydrate content suggesting the notion that melatonin effect on carbohydrate is via H
2S. Taken together, our studyshows the importance of H
2S in melatonin-induced heat stress tolerance through enhancing carbohydrate metabolism under heat stress. It may be emphasized that melatonin modulated carbohydrate metabolism under heat stress to regulate tolerance of heat and caused reduction in oxidative stress and anincrease in antioxidative metabolism via H
2S. Possibly, melatonin functions upstream of H
2S in inducing heat stress signalling response. Such study of regulation of carbohydrate metabolism by H
2S in heat stress is not reported till date and this explores the possibility of H
2S application as heat stress mitigator in plants. In order to make the concept clear, a diagrammatic representation of the effect of melatonin and H
2S on carbohydrate metabolism under heat stress is given (
Figure 9).


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








