
Multi-Omics Research Accelerates the Clarification of the Formation Mechanism and the Influence of Leaf Color Variation in Tea (Camellia sinensis) Plants
 ,
,
Abstract
:1. Introduction
2. Multi-Omics Approaches Further Our Understanding of Leaf Color Variation in Tea
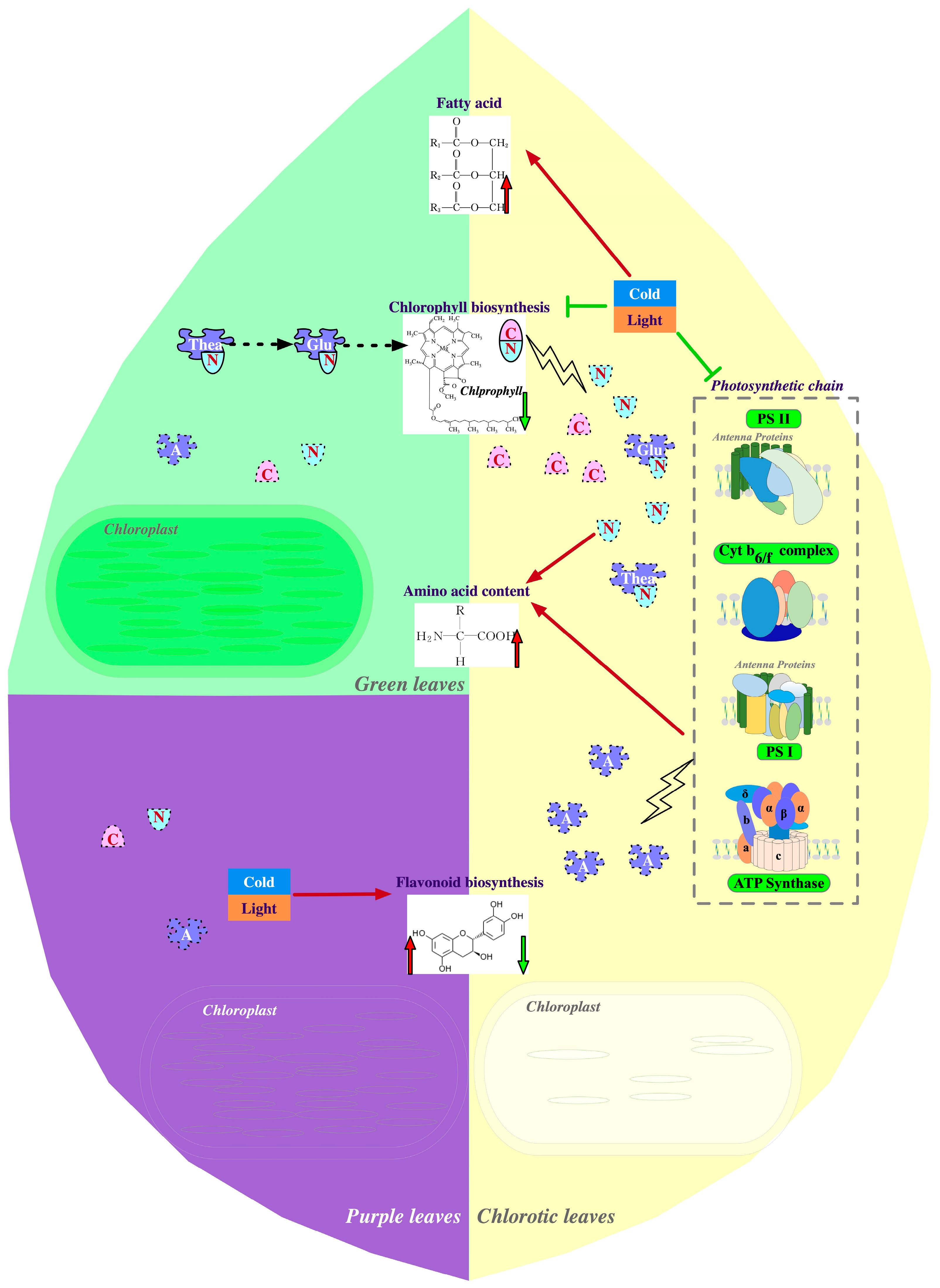
2.1. Physiological Mechanisms of Albinism or Etiolation in Tea
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- ** for free amino acid content using an albino population and SNP markers provides insight into the genetic improvement of tea plants. Hortic. Res. 2022, 9, uhab029. [Google Scholar] [CrossRef]
- Yu, Y.; Kou, X.B.; Gao, R.S.; Chen, X.F.; Zhao, Z.; Mei, H.L.; Li, J.J.; Jeyaraj, A.; Thangaraj, K.; Periakaruppan, R.; et al. Glutamine synthetases play a vital role in high accumulation of theanine in tender shoots of albino tea germplasm “Huabai 1”. J. Agric. Food Chem. 2021, 69, 13904–13915. [Google Scholar] [CrossRef]
- Li, C.F.; Xu, Y.X.; Ma, J.Q.; **, J.Q.; Huang, D.J.; Yao, M.Z.; Ma, C.L.; Chen, L. Biochemical and transcriptomic analyses reveal different metabolite biosynthesis profiles among three color and developmental stages in ‘Anji Baicha’ (Camellia sinensis). BMC Plant Biol. 2016, 16, 195. [Google Scholar] [CrossRef]
- Fu, X.M.; Cheng, S.H.; Liao, Y.Y.; Xu, X.L.; Wang, X.C.; Hao, X.Y.; Xu, P.; Dong, F.; Yang, Z.Y. Characterization of l-theanine hydrolase in vitro and subcellular distribution of its specific product ethylamine in tea (Camellia sinensis). J. Agric. Food Chem. 2020, 68, 10842–10851. [Google Scholar] [CrossRef]
- Liao, H.S.; Chung, Y.H.; Hsieh, M.H. Glutamate: A multifunctional amino acid in plants. Plant Sci. 2022, 318, 111238. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Wang, T.F.; Gan, Q.; Liu, S.; Wang, L.; **, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef] [PubMed]
- Tattini, M.; Galardi, C.; Pinelli, P.; Massai, R.; Remorini, D.; Agati, G. Differential accumulation of flavonoids and hydroxycinnamates in leaves of Ligustrum vulgare under excess light and drought stress. New Phytol. 2004, 163, 547–561. [Google Scholar] [CrossRef] [PubMed]
- Fujii, S.; Wada, H.; Kobayashi, K. Role of Galactolipids in plastid differentiation before and after light exposure. Plants 2019, 8, 357. [Google Scholar] [CrossRef]
- Pye, V.E.; Christensen, C.E.; Dyer, J.H.; Arent, S.; Henriksen, A. Peroxisomal plant 3-ketoacyl-coa thiolase structure and activity are regulated by a sensitive redox switch. J. Biol. Chem. 2010, 285, 24078–24088. [Google Scholar] [CrossRef] [PubMed]
- Arent, S.; Pye, V.E.; Henriksen, A. Structure and function of plant acyl-CoA oxidases. Plant Physiol. Biochem. 2008, 46, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.U. Lipid Metabolism in Plants. Plants 2020, 9, 871. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.W.; Lin, Y.T.; Li, H.M. Increased ratio of galactolipid MGDG: DGDG induces jasmonic acid overproduction and changes chloroplast shape. New Phytol. 2020, 228, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, R.; Wang, G.; Li, M.; Roth, M.; Welti, R.; Wang, X. Differential changes in galactolipid and phospholipid species in soybean leaves and roots under nitrogen deficiency and after nodulation. Phytochemistry 2013, 96, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.Y.; Lam, S.M.; Zuo, J.H.; Yuan, S.Z.; Lv, J.Y.; Shi, J.Y.; Gao, L.P.; Chen, B.; Sui, Y.; Shui, G.H.; et al. Lipidomics reveals the difference of membrane lipid catabolism between chilling injury sensitive and non-sensitive green bell pepper in response to chilling. Postharvest Biol. Technol. 2021, 182, 111714. [Google Scholar] [CrossRef]
- Zhou, Z.; Chen, M.j.; Wu, Q.j.; Zeng, W.; Chen, Z.d.; Sun, W.j. Combined analysis of lipidomics and transcriptomics revealed the key pathways and genes of lipids in light-sensitive albino tea plant (Camellia sinensis cv. Baijiguan). Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Yang, X.Y.; Lu, M.Q.; Wang, Y.F.; Wang, Y.R.; Liu, Z.J.; Chen, S. Response mechanism of plants to drought stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Bhanot, V.; Fadanavis, S.V.; Panwar, J. Revisiting the architecture, biosynthesis and functional aspects of the plant cuticle: There is more scope. Environ. Exp. Bot. 2021, 183, 104364. [Google Scholar] [CrossRef]
- Saini, P.; Bhatia, S.; Mahajan, M.; Kaushik, A.; Sahu, S.K.; Kumar, A.; Satbhai, S.B.; Patel, M.K.; Saxena, S.; Chaurasia, O.P.; et al. ELONGATED HYPOCOTYL5 negatively regulates DECREASE WAX BIOSYNTHESIS to increase survival during UV-B stress. Plant Physiol. 2020, 184, 2091–2106. [Google Scholar] [CrossRef]
- Song, S.S.; Tao, Y.; Gao, L.H.; Liang, H.L.; Tang, D.S.; Lin, J.; Wang, Y.C.; Gmitter, F.G.; Li, C.F. An integrated metabolome and transcriptome analysis reveal the regulation mechanisms of flavonoid biosynthesis in a purple tea plant cultivar. Front. Plant Sci. 2022, 13, 880227. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Chen, X.Y.; Chen, J.H.; Zheng, P.; Liu, S.Q.; Tan, X.D.; Sun, B.M. Virus-induced gene silencing in the tea plant (Camellia sinensis). Plants 2023, 12, 3162. [Google Scholar] [CrossRef] [PubMed]
- Li, G.D.; Li, Y.; Yao, X.Z.; Lu, L.T. Establishment of a virus-induced gene-silencing (VIGS) system in tea plant and its use in the functional analysis of CsTCS1. Int. J. Mol. Sci. 2023, 24, 392. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Tea Varieties | Omics Approaches | Potential Molecular Mechanisms | References |
|---|---|---|---|
| Anjibaicha (Alternative names: Baiye No1, White leaf No.1, and Anji white 1) | Succinyl-proteome | Photosynthetic chain: The succinylation levels of PsbS and light-harvesting complex LHCA4 are down-regulated; the succinylation level of LHCB4 is up-regulated. | [46] |
| Proteome and acetyl-proteome | Photosynthetic chain: lower abundance of LHCB1, LHCB2, LHCB3, LHCB4, LHCB5, LHCB7, LHCA1, LHCA2, LHCA3, LHCA4, PsbC, PsbD, PsbO, PsbP, PsbQ, PsbR, PsbS, Psb27, PsaE, PsaG, PsaL, PsaN, PetA, PetC, PetE, PetF, PetH, ATPA, ATPB, ATPD, ATPE, ATPG; a lower acetylation level of LHCA1. | [8] | |
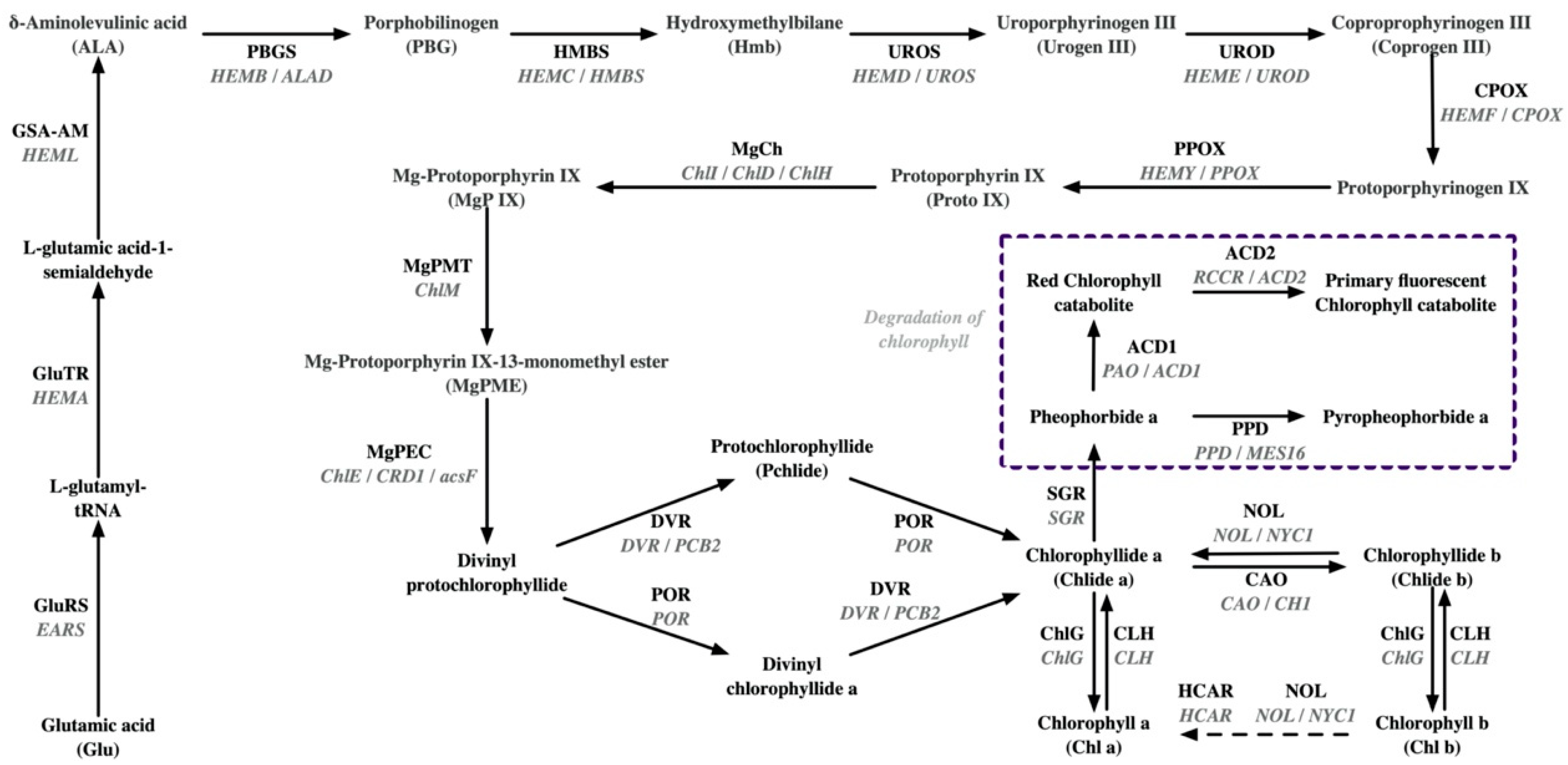
| Pangenome | Chlorophyll synthesis: The GluRS/EARS gene in Anjibaicha showed a loss of anti-codon recognition domains; SVs were revealed in ChlD. Chlorophyll degradation: 1 bp deletion is revealed in NOL/NYC1. | [24] | |
| Whole-transcriptome | Chlorophyll synthesis: down-regulated expression of POR (two alleles), CLH1, PBGD/HMBS/HEMC; up-regulated expression of COX15. Photosynthetic chain: down-regulated expression of CAB7, CAB21, LHCA4, CAB40 (three alleles), CAB13, LHCB5, PetA (two alleles), PsbA (TEA_001460), PsaB, PsbP1, ATPA, PsaH, ATPI, PetB, Psb28; up-regulated expression of PsbA (TEA_001460). | [35] | |
| Huang**ya | Proteome | Chlorophyll synthesis: higher abundance of GluRS/EARS, MgCh/ChlH, and HMBS; lower abundance of POR. Photosynthetic chain: lower abundance of Photosystem Q(B) protein. | [36] |
| Metabolome and proteome | Chlorophyll synthesis: higher abundance of GluRS/EARS and UROD/HEME; lower abundance of POR and CAO. Chlorophyll degradation: higher abundance of PAO/ACD1 Photosynthetic chain: lower abundance of LHCA1, LHCA3, LHCB1, LHCB2, LHCB3, LHCB4, LHCB5, LHCB6, PsaD, PsaF, PsaL, PsaN, PsbA, PsbD, PsbE, PsbO, PsbP, PsbQ; higher abundance of PsbS, FNR, ATPB. Carotenoid synthesis: lower abundance of ZEP; higher abundance of PSY. | [9] | |
| Pangenome | Chlorophyll synthesis: The GluRS/EARS gene in Huang**ya showed a loss of anti-codon recognition domains, which may inhibit chlorophyll synthesis; SVs were detected in CAO, CHLP, and GluTR. Carotenoid synthesis: mutated amino acids in CYP97A3HJY and elevated expression of the CYP97A3HJY allele. | [24] | |
| Transcriptome | Carotenoid synthesis: down-regulated expression of PSY, PDS, ZDS, LCYE, LCYB, CHY, ZEP, and VDE. | [47] | |
| Baijiguan | BSR-seq | Photosynthetic chain: co-down-regulated expression of LHCA3, three LHCB1 alleles (TEA001863, TEA001868, TEA030368), two LHCB3 alleles (TEA017256, TEA021966), and LHCB4 in bulked groups and parents. | [29] |
| Genome: Genoty** by sequencing and BSA-seq | Chlorophyll synthesis: a non-synonymous polymorphism (G1199A) in the magnesium chelatase I subunit (CsChlI). | [38] | |
| HY1 | Proteome | Photosynthetic chain: lower abundance of LHCA3, LHCA4, LHCB1, LHCB2, LHCB6, PsbC, PsBO, PsbS, PsaB, PsaC, PsaD, PsaF, PetA, PetH. | [17] |
| HY2 | Proteome | Photosynthetic chain: lower abundance of LHCB2, PsbA, PsbD, PsbC, PsbB, PsbQ, PsaA, PsaB, PsaD, PsaH, PetB, PetA, beta F-type ATPase. | [17] |
| HY | Transcriptome | Chlorophyll degradation: the activation of SGR and CLH. | [43] |
| **angfeihuangye | Transcriptome, translatome, and metabolome | Chlorophyll synthesis: up-regulation of HY5 in EL inhibited the expression of GluTR/HEMA and POR. | [39] |
| Huabai 1 | Transcriptome | Photosynthetic chain: down-regulated expression of light-harvesting complex II (LHCII) chlorophyll-a/b-binding protein. Chlorophyll degradation: up-regulated expression of SGR. | [12] |
| Albinistic branch of Huangshan | Transcriptome | Chlorophyll synthesis: down-regulated expression of four CHLP alleles (TEA027589, TEA019124, BGI_novel_G007262, TEA016514) and one POR allele (TEA014780); up-regulated expression of one CHLP allele (TEA009538), one POR allele (TEA027994), and one CLH allele (TEA027808). Photosynthetic chain: down-regulated expression of three LHCB1 alleles (TEA019232, TEA030366, TEA030368), PsbC, five PsbB alleles (TEA028468, TEA011113, TEA032780, TEA018797, BGI_novel_G013475), PsbP, Psb28, PsaA, five ATPD alleles (TEA030038, TEA004696, TEA002611, BGI_novel_G006800, BGI_novel_004911), two ATPA alleles (TEA019276, BGI_novel_G009498), ATPE. | [48] |
| Huang**ju | Transcriptome | Chlorophyll synthesis: up-regulated expression of POR. Photosynthetic chain: down-regulated expression of LHCA2, LHCA4, LHCB1, LHCB2, LHCB6. | [13] |
| Yanlingyinbiancha | Transcriptome | Chlorophyll synthesis: down-regulated expression of UROS/HEMD, PPOX, ChlH/GUN5, MgPEC/CRD1, DVR/PCB2, and CAO. Chlorophyll degradation: down-regulated expression of NOL/NYC1, HCAR, CLH1, and ACD2. Photosynthetic chain: fifty-five DEGs involved in photosynthetic complexes were found to be down-regulated. Carotenoid synthesis: down-regulated expression of Z-ISO, ZDS, ZEP, LUT2, NCED4. | [18] |
| Yanling Huayecha | Transcriptome | Chlorophyll synthesis: down-regulated expression of PPOX. Photosynthetic chain: down-regulated expression of LHCB6 and FdC2. Thylakoid membrane structure: down-regulated expression of SCY1. | [40] |
| Menghai Huangye | Transcriptome | Chlorophyll synthesis: four genes related to chlorophyll synthesis (HEME2 and POR). Photosynthetic chain: ten genes related to photosynthesis (LHCA and LHCB) are down-regulated. | [41] |
| Zhonghuang 3 | Transcriptome | Chlorophyll metabolism: down-regulated expression of GluTR/HEMA3 and CLH4. | [49] |
| Zhonghuang 3 | Transcriptome | Chlorophyll synthesis: down-regulated expression of GluTR/HEMA, GSA-AM/HEML, UROD/HEME, HEMF/CPOX, DVR (CSS0009780), and CHLP; up-regulated expression of PBGS/HEMB, DVR (CSS0011936), and CLH. Chlorophyll degradation: down-regulated expression of NOL/NYC1 (CSS0031926) and SGR (CSS0030812); up-regulated expression of NOL/NYC1 (CSS0015127), SGR (CSS0050352), and SGRL (CSS0004139 and CSS0036450). Carotenoid synthesis: up-regulated expression of Z-ISO, CRTISO (CSS0027469, CSS0033902, and CSS0044870), NCED1, NCED2; down-regulated expression of LCYB and NXS. | [14] |
| Zhonghuang 2 | Transcriptome | Transcripts encoding enzymes such as those functioning in early enzymatic steps, from the formation of glutamate 1-semialdehyde to protoporphyrin IX, showed lower levels. Critical enzymes for converting Mg-protoporphyrin IX into chlorophyll were also inhibited. | [42] |
| Koganemidori | Transcriptome | Chlorophyll synthesis: down-regulated expression of POR, CAO, and ChlG. Chlorophyll degradation: up-regulated expression of CLH. Transcriptional regulation: two homologs of GLK were significantly down-regulated. | [50] |
| Huang**shuixian | Transcriptome and metabolome | Chlorophyll degradation: down-regulated expression of SGR. Carotenoid synthesis: the expression of DXS and GGPPS was significantly down-regulated. Transcriptional regulation: PIFs related to chlorophyll biosynthesis were significantly suppressed. | [16] |
| Huangyu | Transcriptome and metabolome | Chlorophyll synthesis: down-regulated expression of UROD/HEME, MgCh/ChlH, and CAO. Chlorophyll degradation: up-regulated expression of CLH. Photosynthetic chain: down-regulated expression of three LHCII genes (CSS0013089, CSS0017825, and CSS0039893) | [30] |
| Huangkui | Transcriptome | Transcriptional regulation: the transcriptional expression of CsRVE1 increased during seasonal greening and was tightly correlated with increases in the expression of genes involved in light harvesting (LHCB) and chlorophyll biosynthesis (MgCh/ChlH, GluTR/HEMA1, and CAO). | [51] |
| Fuhuang 1 | Transcriptome | Chlorophyll synthesis: down-regulated expression of CAO. Chlorophyll degradation: down-regulated expression of NOL/NYC1 and SGR. Photosynthetic chain: down-regulated expression of LHCA2, LHCA4, LHCB1, LHCB3. Carotenoid synthesis: down-regulated expression of LCYE, ZEP, NCED; up-regulated expression of PSY, PDS, VDE. | [52] |
| Fuhuang 2 | Transcriptome | Chlorophyll synthesis: down-regulated expression of GluTR/HEMA. Chlorophyll degradation: down-regulated expression of CLH. Photosynthetic chain: down-regulated expression of PsbB, PetC, ATPF1B, LCHBs, LCHAs. Carotenoid synthesis: down-regulated expression of NCED. | [53] |
| Tea Varieties | Sampling Location | Measurement Technique | Anthocyanin Composition | References |
|---|---|---|---|---|
| Zijuan | Tea garden of South China Agricultural University, Guangzhou, China | HPLC | Major anthocyanin compositions: cyanidin-3-O-galactoside and delphinidin-3-O-galactoside. | [77] |
| Tea garden of the Institute of Tea Science, Yunnan Province Academy of Agricultural Sciences (Menghai, China) | Non-targeted metabolomics approach: UHPLC– Orbitrap–MS/MS | Cyanidin 3-diglucoside 5-glucoside, cyanidin 3-O-(6-O-p-coumaroyl) glucoside, cyanidin 3-sambubioside, cyanidin 3-(6″-acetylglucoside)-5-glucoside, delphinidin 3-(6-p-coumaroyl) galactoside, delphinidin-3-O-arabinoside, pelargonidin 3-sophoroside 5-glucoside, pelargonidin 3-coumarylglucoside-5-acetylglucoside, pelargonidin 3-rhamnoside 5-glucoside; compared with Yunkang, the contents of cyanidin 3-diglucoside 5-glucoside and pelargonidin 3-sophoroside 5-glucoside are most increased in Zijuan. | [86] | |
| Pu’er City Institute of Tea Science, Yunnan Province | UPLC–ESI–MS/MS metabolomic analysis | Specific metabolites: petunidin 3-O-glucoside, peonidin 3-O-glucoside chloride, peonidin 3-O-glucoside, peonidin O-hexoside, malvidin 3-O-glucoside (oenin), petunidin 3,5-O-diglucoside. Marker metabolites: cyanidin 3-O-galactoside, cyanidin 3-O-glucoside (Kuromanin), delphinidin 3-O-glucoside (Mirtillin), pelargonidin 3-O-glucoside | [87] | |
| Changsha, Hunan, China | UPLC–ESI–MS/MS metabolomic analysis | Major anthocyanin compositions: cyanidin-3-ogalactoside, delphinidin-3-O-galactoside, and petunidin-3-O-galactoside | [88] | |
| Dechang Fabrication Base of Shucheng County in Anhui Province, China | LC−TOF–MS | Cyanidin-3-O-galactoside, Cyanidin 3-O-(6-O-p-coumaroyl) galactoside, Delphinidin 3-O-(6-O-p-coumaroyl) galactoside, Delphinidin-3-O-galactoside. | [89] | |
| Zijuan Ziyan and Chuanzi (ZZ) | Muchuan County, Sichuan Province, China | Targeted UPLC– ESI–MS/MS analysis | A total of 22 anthocyanins with a content ≥1 μg/g (DW) were detected in Chuanzi, Ziyan, and/or Zijuan and these included 6 cyanidins, 7 delphinidins, 5 pelargonidins, 2 peonidins, and 2 petunidins. In addition, 23 anthocyanins with a concentration of <1 μg/g were also detected. | [82] |
| Ziyan | Planted in plastic pots | HPLC | Delphinidin, cyanidin, and pelargonidin. | [90] |
| Hongyecha, Zijuan, 9803, Hongyafoshou | Changsha, Hunan, China | UPLC–DAD–QTOF–MS | Cyanidin-(E)-p-coumaroylgalactoside, cyanidin-3-O-galactoside, delphinidin-3-O-galactoside, delphinidin-(Z)-p-coumaroylgalactoside, delphinidin-(E)-p-coumaroylgalactoside, pelargonidin-O-hexose, and pelargonidin-O-dihexose. | [91] |
| **mingzao | Tea plantation of Wuqu in Fuan City, Fujian Province, China | Widely targeted metabolomics: UPLC–ESI–MS/MS | Cyanidin 3-O-glucoside, cyanidin 3-O-galactoside, cyanidin 3-rutinoside, cyanidin chloride, delphinidin 3-O-glucoside, peonidin 3-O-glucoside chloride (most affected). | [83] |
| Zikui | South Campus of Guizhou University, Huaxi District, Guiyang City, Guizhou Province, China | ESI–QTRAP–MS/MS | Cyanidin 3-O-galactoside, cyanidin 3-O-glucosid, petunidin 3-O-glucoside. | [84] |
| Long**g43 | Tea Research Institute, Chinese Academy of Agricultural Sciences, Hangzhou, China | LC–MS/MS | Delphinidin-hexose-coumaroyl showed the greatest increase. | [80] |
| TRFK 306 | Tea Research Institute (TRI), Kericho County, Kenya | HPLC | Malvidin 3-glucoside, peonidin 3-glucoside, pelargonidin 3,5-O-diglucoside, cyanidin 3-O-glucoside, cyanidin 3-O-galactoside, cyanidin 3-O-rutinoside. | [81] |
| 9 tea cultivars possessing purple leaves | Wuxi Institute of Tea Varieties in Wuxi City, Jiangsu Province, China | Widely targeted metabolomics: UPLC–ESI–MS/ MS | Thirty-three anthocyanins were identified, and delphinidin 3-O-galactoside and cyanidin 3-O-galactoside were found to be the most abundant in PTLs. | [85] |
| Unknown | Experimental tea farm (IHBT-269) of CSIR—Institute of Himalayan Bioresource Technology, HP, India | UHPLC | 3-O-alpha-l-arabinopyranosylproantho cyanidin A5′ and 3,3′-Di-O-galloylprocyanidin B. | [92] |
| Tea Varieties | Omics Approaches | Potential Molecular Mechanisms | References |
|---|---|---|---|
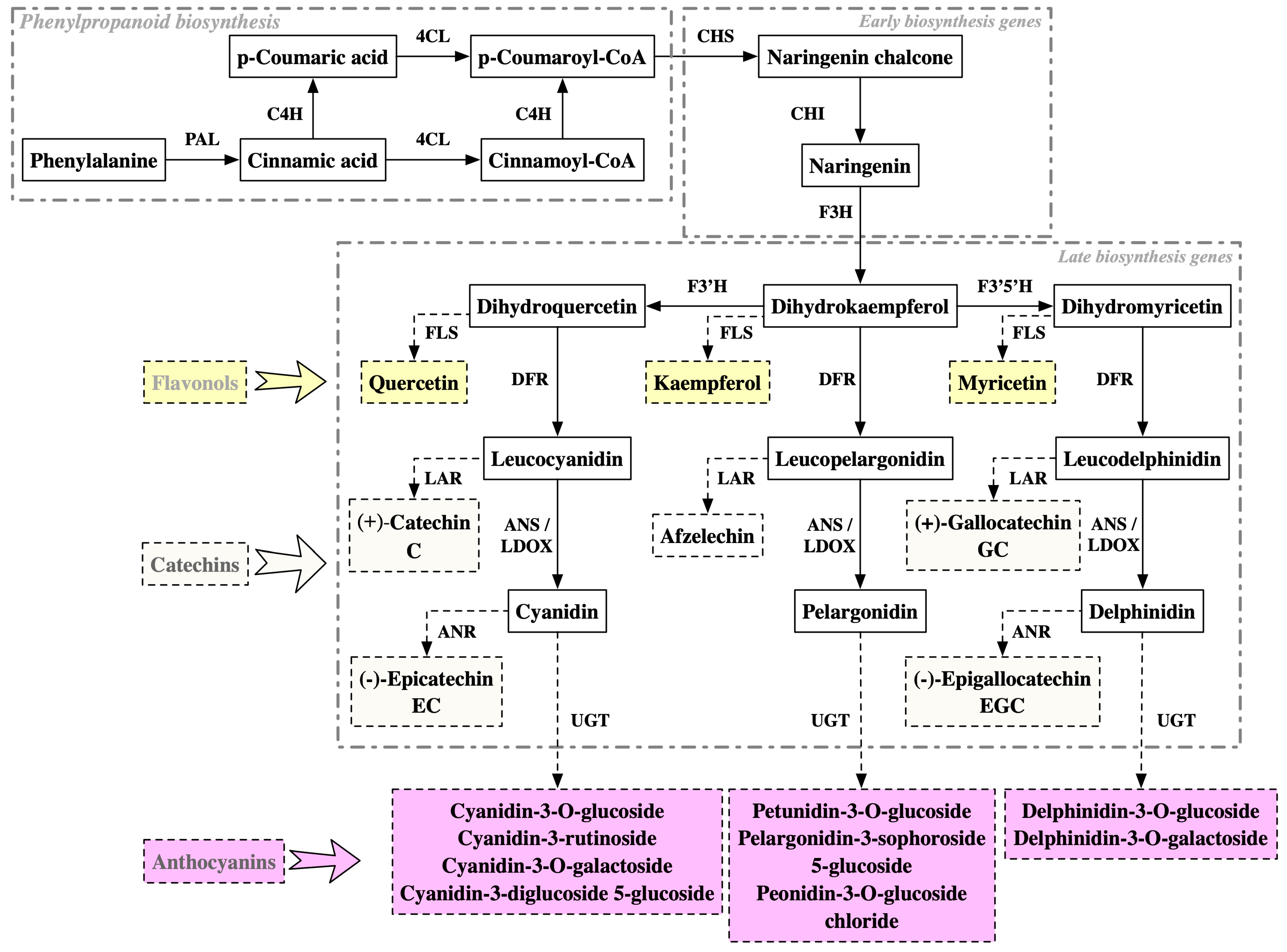
| Zijuan | Transcriptome | Transcriptional regulation: Activation of the R2R3-MYB transcription factor (TF) anthocyanin1 (CsAN1) and the bHLH TF CsGL3; CsAN1 interacts with bHLH TFs (CsGL3 and CsEGL3) and recruits a WD-repeat protein CsTTG1 to form the MYB-bHLH-WDR (MBW) complex that regulates anthocyanin accumulation. Late biosynthetic genes (LBGs): activation of CsF3′H, CsF3′5′H, CsDFR1, CsDFR2, CsANS1/LDOX1, CsANS2/LDOX2, and CsANS3/LDOX3. Metabolic substrate competition: activation of CsLAR1, CsLAR2, and CsLAR3, which encode enzymes for catechin biosynthesis, was highly expressed in red foliage. | [77] |
| Transcriptome, proteome | Phenylpropanoid metabolism: significantly increased expression of three PALs (CSA016076, 022024, 022025); significantly decreased expression of 4CL (CSA001434). Early biosynthesis genes (EBGs): significantly increased expression of CHS (CSA029775); significantly decreased expression of CHI (CSA008261). LBGs: significantly increased expression of two DFRs (CSA003949, XLOC_010242), one ANS/LDOX (CSA011508), six UGT75L12/13 (CSA005544, 005545, 010001, 036671, 036672, 029026), and two UGT94P1 (CSA007394, 008750); significantly decreased expression of F3′5′H (CSA031792), ANS/LDOX (CSA035767), two UGT75L12s (CSA008693, 028873), and two UGT94P1s (CSA005965, 026000). Metabolic substrate competition: significantly increased expression of two LARs (CSA014943, XLOC_016774). | [87] | |
| Transcriptome | Phenylpropanoid metabolism: activation of C4H. LBGs: activation of ANS/LDOX, UGT. Chlorophyll degradation: activation of CLH1. | [97] | |
| Full-length transcriptome | Alternative splicing (AS) events identified in transcriptional regulation (MYB113-1), phenylpropanoid metabolism (C4H1, PAL2), LBGs (UDP75L122), and metabolic substrate competition (FLS1). | [98] | |
| Proteome | EBGs: increased abundance of CHS and CHI. LBGs: increased abundance of DFR, ANS/LDOX, and UGT. Anthocyanin transportation: increased abundance of ABC transporter B8. | [20] | |
| Transcriptome | Transcriptional regulation: Most of the members belonging to the MYB, WRKY, AP2, GRF, bZIP, and MYC groups had a higher expression in Zijuan. LBGs: significantly increased expression of F3′5′H (CSS0022212.1), ANS/LDOX (CSS0010687.1), 3GT (anthocyanidin 3-O-glucosyltransferase, CSS0024320.1), 3AT (cyanidin-3-O-glucoside 6″-O-acyltransferase, CSS0015285.1). Metabolic substrate competition: significantly decreased expression of LAR (CSS0009063.1). Anthocyanin degradation: polyphenol oxidase (PPO, CSS0002951.1), showed negative correlation with the three anthocyanins, especially delphinidin and delargonidin. | [89] | |
| Chuanzi (ZZ) | Transcriptome | Transcriptional regulation: significantly increased expression of the well-known MYB transcription factor CsAN1/CsMYB75 (CSS0030514). LBGs: significantly increased expression of CsANSs/LDOXs (CSS0010687, CSS0018498 and CSS0046216), CsUGT94P1 (CSS0011196), and the anthocyanin O-methyltransferase gene (CsAOMT, CSS0015915). Anthocyanin transportation: significantly increased expression of CsGSTF1 (CSS0022086) and three other GST candidate genes (CSS0031248, CSS0026690, and CSS0018634) tightly linked to CsGSTF1. Metabolic substrate competition: down-regulated expression of LARs (CSS0028235 and CSS0009063) and ANRs (CSS0005927, and CSS0033195). | [82] |
| Zijuan, **guanyin and **mingzao | Pangenome | Read depth of the LTR insertion region in the promoter of CsMYB114 among a set of representative purple-leaf cultivars (‘ZJ’, ‘JMZ’, and ‘JGY’) and tea cultivars with green leaves (‘FDDB’, ‘BHZ’, and ‘GH3H’) | [24] |
| Ziyan | Transcriptome | Transcriptional regulation: UV-A induces the expression of the regulatory gene TT8; UV-AB induces the expression of the regulatory genes EGL1 and TT2. LBGs: UV-A induces the expression of F3H, F3′5′H, DFR, and ANS/LDOX; UV-AB induces the expression of F3′5′H, DFR, ANS/LDOX, and UGT. Metabolic substrate competition: UV radiation repressed the expression levels of LAR, ANR, and FLS, resulting in reduced ANR activity and a metabolic flux shift towards anthocyanin biosynthesis. | [90] |
| Wuyiqizhong18 | cDNA-AFLP | EBGs: increased expression of CHS. LBGs: increased expression of AT (TDF #3341_2f) and UGT (TDF #2421_1d and TDF #2411_1f). | [79] |
| Proteome | EBGs: increased abundance of CHS and CHI. Metabolic substrate competition: increased abundance of FLS. | [99] | |
| **mingzao | Transcriptome | Phenylpropanoid metabolism: activation of PAL, C4H, and 4CL. LBGs: activation of DFR, ANS/LDOX, and UGT (TEA004632 and TEA004632) genes. | [83] |
| Long**g43 | Transcriptome | Transcriptional regulation: activation of MYB75. LBGs: activation of ANS/LDOX and 3-GT. Anthocyanin transportation: activation of genes involved in anthocyanin transportation (GST, glutathione S-transferase). | [96] |
| Transcriptome | Phenylpropanoid metabolism: activation of PAL and C4H by high temperature and/or light levels in summer. EBGs: activation of CHI and CHS by high temperature and/or light levels in summer. LBGs: activation of ANR, ANS/LDOX, and DFR by high temperature and/or light levels in summer. Metabolic substrate competition: activation of FLS and LAR by high temperature and/or light levels in summer. | [80] | |
| TRFK 306 | Transcriptome | Transcriptional regulation: transcripts encoding pathway regulators of the MYB–bHLH–WD40 (MBW) complex were repressed, possibly contributing to the suppression of late biosynthetic genes of the pathway during the dry season. Anthocyanin transportation: suppression of anthocyanin transport genes could be linked to reduced accumulation of anthocyanin in the vacuole during the dry season. | [81] |
| Zikui | Transcriptome | Transcriptional regulation: CsMYB90 showed strong correlations with petunidin 3-O-glucoside, cyanidin 3-O-galactoside, and cyanidin 3-O-glucosid. LBGs: activation of two F3′H genes and two ANS/LDOX genes. Anthocyanin degradation: three negatively correlated PPO (polyphenol oxidase) genes with anthocyanin accumulation. | [84] |
| Hongyecha, Zijuan, 9803, Hongyafoshou | Transcriptome | Phenylpropanoid metabolism: activation of 4CL. LBGs: activation of ANS/LDOX and UGT. | [88] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Y.-G.; Zhao, T.-T.; **ang, Q.-Z.; Han, X.-Y.; Yang, S.-S.; Zhang, L.-X.; Ren, L.-J. Multi-Omics Research Accelerates the Clarification of the Formation Mechanism and the Influence of Leaf Color Variation in Tea (Camellia sinensis) Plants. Plants 2024, 13, 426. https://doi.org/10.3390/plants13030426
Fan Y-G, Zhao T-T, **ang Q-Z, Han X-Y, Yang S-S, Zhang L-X, Ren L-J. Multi-Omics Research Accelerates the Clarification of the Formation Mechanism and the Influence of Leaf Color Variation in Tea (Camellia sinensis) Plants. Plants. 2024; 13(3):426. https://doi.org/10.3390/plants13030426
Chicago/Turabian StyleFan, Yan-Gen, Ting-Ting Zhao, Qin-Zeng **ang, **ao-Yang Han, Shu-Sen Yang, Li-**a Zhang, and Li-Jun Ren. 2024. "Multi-Omics Research Accelerates the Clarification of the Formation Mechanism and the Influence of Leaf Color Variation in Tea (Camellia sinensis) Plants" Plants 13, no. 3: 426. https://doi.org/10.3390/plants13030426