
miR-125 in Breast Cancer Etiopathogenesis: An Emerging Role as a Biomarker in Differential Diagnosis, Regenerative Medicine, and the Challenges of Personalized Medicine

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Clinical Features of BC
2.1. Main Risk Factors of BC
2.2. BC Characterization and Patient Management
3. Epigenetics of BC and the Role of miR-125
3.1. microRNA Nomenclature
3.2. Role of miR in BC
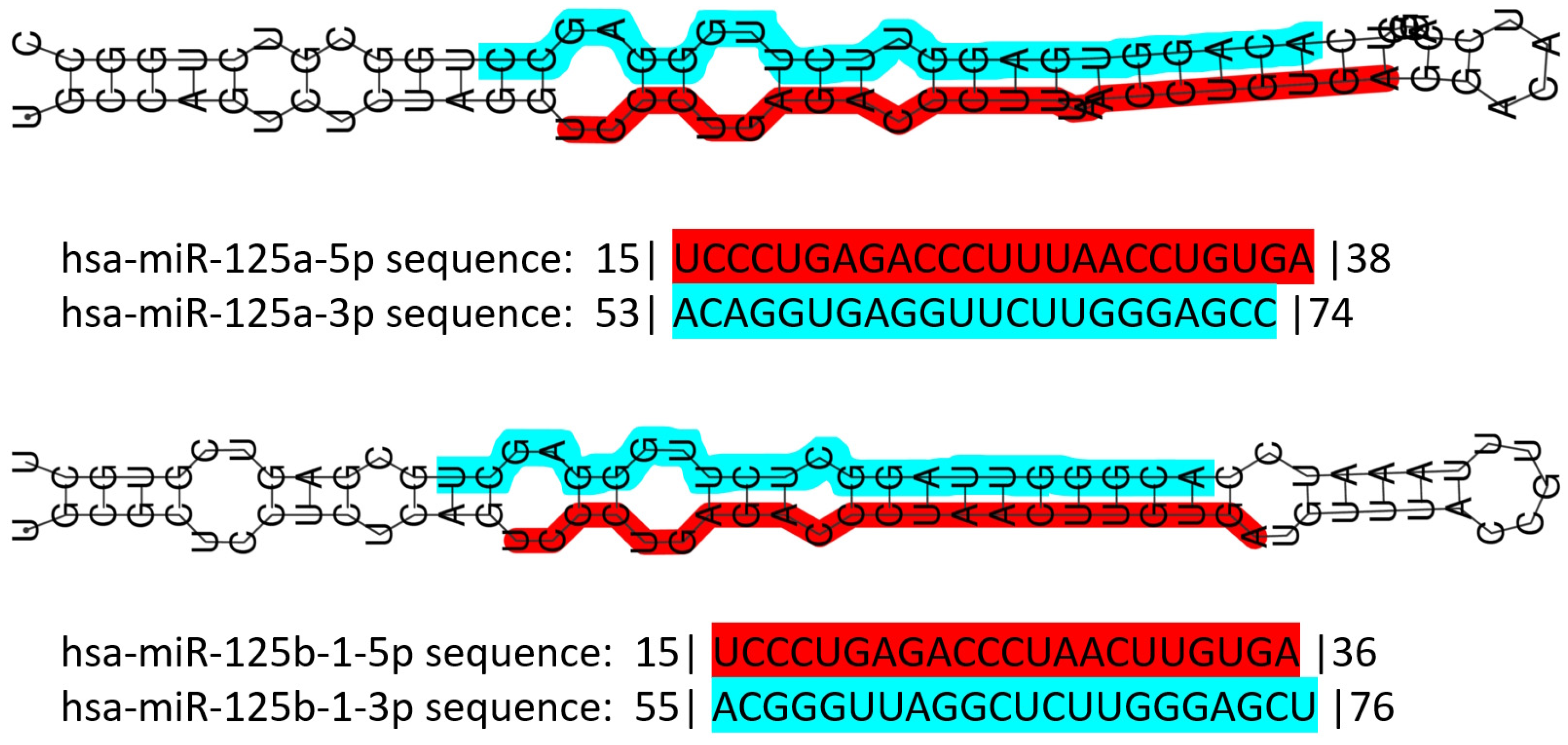
3.3. The miR-125 Family: Molecular Organization and Roles in Human Pathology
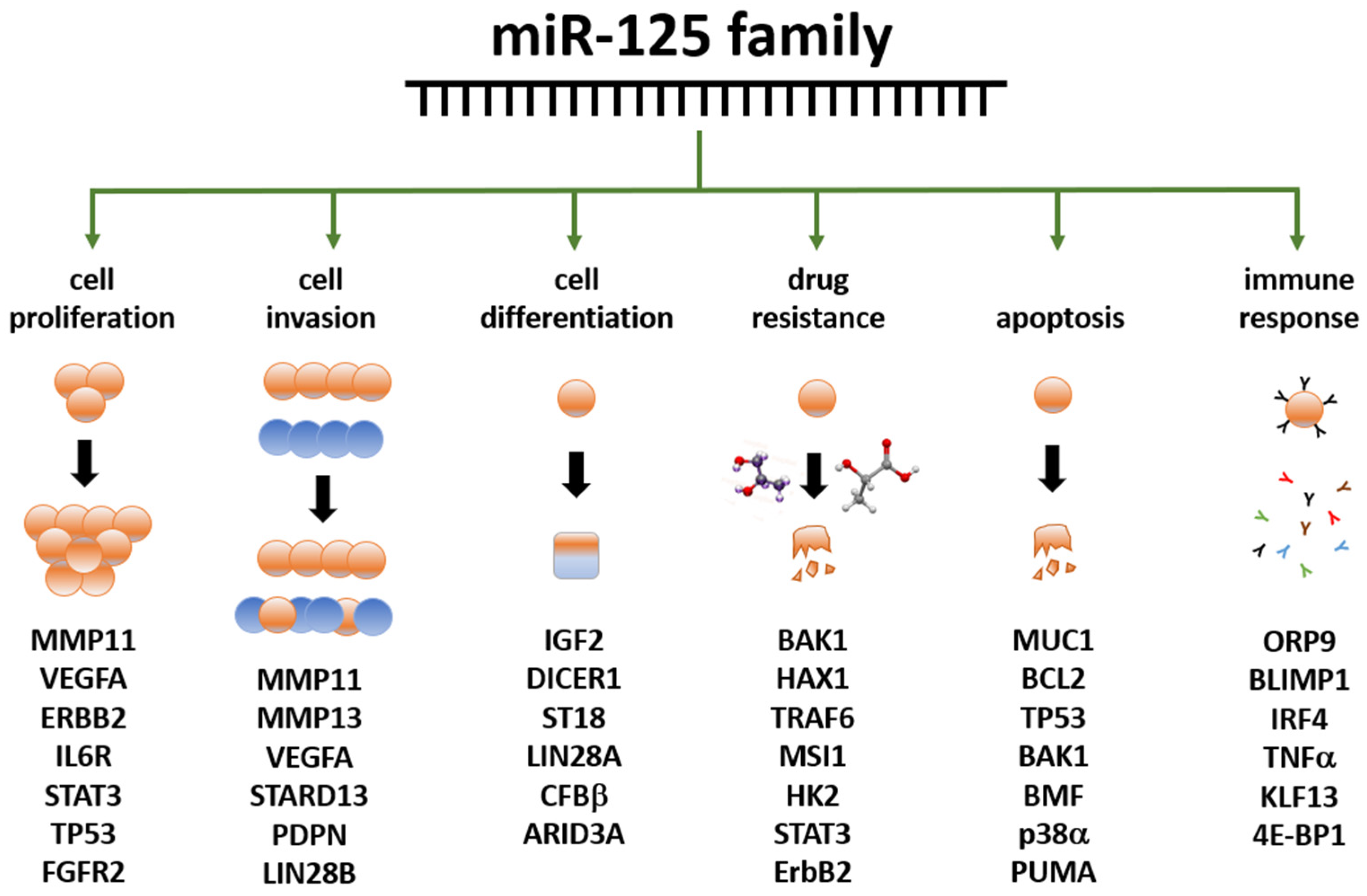
3.4. miR-125 and Cancer
3.5. Role of miR-125 in BC
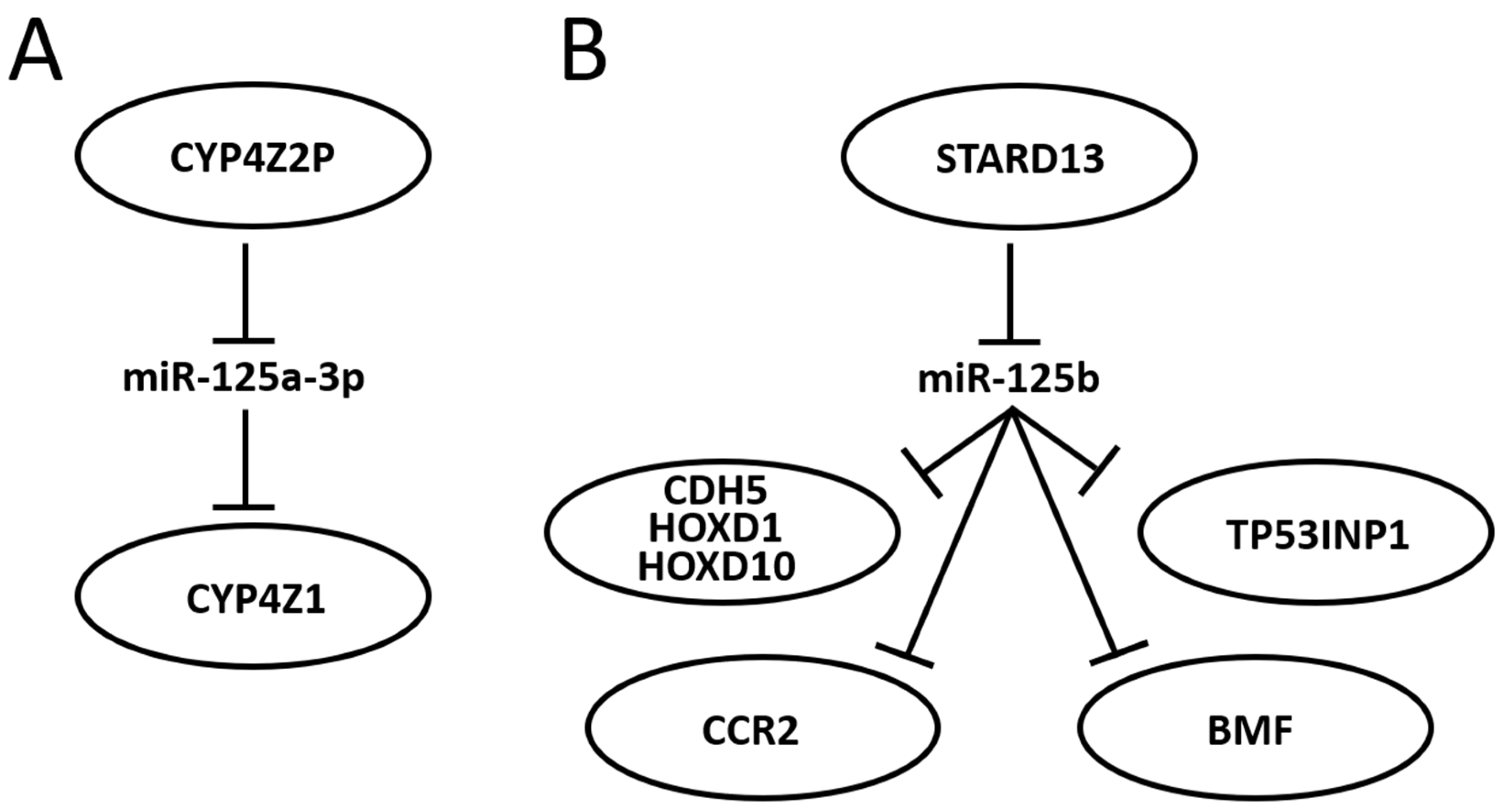
3.6. Further Mining miR-125 Function in BC: Competing Endogenous RNA Networks (ceRNET)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer Statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Winters, S.; Martin, C.; Murphy, D.; Shokar, N.K. Breast Cancer Epidemiology, Prevention, and Screening. Prog. Mol. Biol. Transl. Sci. 2017, 151, 1–32. [Google Scholar] [PubMed]
- Arnold, M.; Morgan, E.; Rumgay, H.; Mafra, A.; Singh, D.; Laversanne, M.; Vignat, J.; Gralow, J.R.; Cardoso, F.; Siesling, S.; et al. Current and Future Burden of Breast Cancer: Global Statistics for 2020 and 2040. Breast 2022, 66, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R. Global, Regional, National Burden of Breast Cancer in 185 Countries: Evidence from GLOBOCAN 2018. Breast Cancer Res. Treat. 2021, 187, 557–567. [Google Scholar] [CrossRef]
- Jung, S.; Wang, M.; Anderson, K.; Baglietto, L.; Bergkvist, L.; Bernstein, L.; van den Brandt, P.A.; Brinton, L.; Buring, J.E.; Heather Eliassen, A.; et al. Alcohol Consumption and Breast Cancer Risk by Estrogen Receptor Status: In a Pooled Analysis of 20 Studies. Int. J. Epidemiol. 2016, 45, 916–928. [Google Scholar] [CrossRef]
- Key, T. Sex Hormones and Risk of Breast Cancer in Premenopausal Women: A Collaborative Reanalysis of Individual Participant Data from Seven Prospective Studies. Lancet Oncol. 2013, 14, 1009–1019. [Google Scholar] [CrossRef]
- Coughlin, S.S. Epidemiology of Breast Cancer in Women. Adv. Exp. Med. Biol. 2019, 1152, 9–29. [Google Scholar] [PubMed]
- Galati, F.; Magri, V.; Arias-Cadena, P.A.; Moffa, G.; Rizzo, V.; Pasculli, M.; Botticelli, A.; Pediconi, F. Pregnancy-Associated Breast Cancer: A Diagnostic and Therapeutic Challenge. Diagnostics 2023, 13, 604. [Google Scholar] [CrossRef]
- Bodewes, F.T.H.; van Asselt, A.A.; Dorrius, M.D.; Greuter, M.J.W.; de Bock, G.H. Mammographic Breast Density and the Risk of Breast Cancer: A Systematic Review and Meta-Analysis. Breast 2022, 66, 62–68. [Google Scholar] [CrossRef]
- Majeed, W.; Aslam, B.; Javed, I.; Khaliq, T.; Muhammad, F.; Ali, A.; Raza, A. Breast Cancer: Major Risk Factors and Recent Developments in Treatment. Asian Pac. J. Cancer Prev. 2014, 15, 3353–3358. [Google Scholar] [CrossRef]
- Petrucelli, N.; Daly, M.B.; Pal, T. BRCA1- and BRCA2-Associated Hereditary Breast and Ovarian Cancer. 4 September 1998 [Updated 21 September 2023]. In GeneReviews® [Internet]; Adam, M.P., Feldman, J., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Gripp, K.W., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993–2024; Available online: https://www.ncbi.nlm.nih.gov/books/NBK1247/ (accessed on 15 October 2023).
- Carbognin, L.; Miglietta, F.; Paris, I.; Dieci, M.V. Prognostic and Predictive Implications of PTEN in Breast Cancer: Unfulfilled Promises but Intriguing Perspectives. Cancers 2019, 11, 1401. [Google Scholar] [CrossRef] [PubMed]
- Shahbandi, A.; Nguyen, H.D.; Jackson, J.G. TP53 Mutations and Outcomes in Breast Cancer: Reading beyond the Headlines. Trends Cancer 2020, 6, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Corso, G.; Veronesi, P.; Sacchini, V.; Galimberti, V. Prognosis and Outcome in CDH1-Mutant Lobular Breast Cancer. Eur. J. Cancer Prev. 2018, 27, 237–238. [Google Scholar] [CrossRef] [PubMed]
- Beggs, A.D.; Latchford, A.R.; Vasen, H.F.A.; Moslein, G.; Alonso, A.; Aretz, S.; Bertario, L.; Blanco, I.; Bülow, S.; Burn, J.; et al. Peutz–Jeghers Syndrome: A Systematic Review and Recommendations for Management. Gut 2010, 59, 975–986. [Google Scholar] [CrossRef] [PubMed]
- Apostolou, P.; Papasotiriou, I. Current Perspectives on CHEK2 Mutations in Breast Cancer. Breast Cancer Targets Ther. 2017, 9, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Nepomuceno, T.C.; Carvalho, M.A.; Rodrigue, A.; Simard, J.; Masson, J.Y.; Monteiro, A.N.A. PALB2 Variants: Protein Domains and Cancer Susceptibility. Trends Cancer 2021, 7, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Stucci, L.S.; Internò, V.; Tucci, M.; Perrone, M.; Mannavola, F.; Palmirotta, R.; Porta, C. The ATM Gene in Breast Cancer: Its Relevance in Clinical Practice. Genes 2021, 12, 727. [Google Scholar] [CrossRef]
- Li, N.; McInerny, S.; Zethoven, M.; Cheasley, D.; Lim, B.W.X.; Rowley, S.M.; Devereux, L.; Grewal, N.; Ahmadloo, S.; Byrne, D.; et al. Combined Tumor Sequencing and Case-Control Analyses of RAD51C in Breast Cancer. J. Natl. Cancer Inst. 2019, 111, 1332–1338. [Google Scholar] [CrossRef]
- Chen, X.; Li, Y.; Ouyang, T.; Li, J.; Wang, T.; Fan, Z.; Fan, T.; Lin, B.; ** Review. Cureus 2023, 15, e44061. [Google Scholar] [CrossRef]
- Abdul Halim, A.A.; Andrew, A.M.; Mohd Yasin, M.N.; Abd Rahman, M.A.; Jusoh, M.; Veeraperumal, V.; Rahim, H.A.; Illahi, U.; Abdul Karim, M.K.; Scavino, E. Existing and Emerging Breast Cancer Detection Technologies and Its Challenges: A Review. Appl. Sci. 2021, 11, 10753. [Google Scholar] [CrossRef]
- Iranmakani, S.; Mortezazadeh, T.; Sajadian, F.; Ghaziani, M.F.; Ghafari, A.; Khezerloo, D.; Musa, A.E. A Review of Various Modalities in Breast Imaging: Technical Aspects and Clinical Outcomes. Egypt. J. Radiol. Nucl. Med. 2020, 51, 57. [Google Scholar] [CrossRef]
- Veronesi, U.; Viale, G.; Rotmensz, N.; Goldhirsch, A. Rethinking TNM: Breast Cancer TNM Classification for Treatment Decision-Making and Research. Breast 2006, 15, 3–8. [Google Scholar] [CrossRef]
- Eliyatkin, N.; Yalcin, E.; Zengel, B.; Aktaş, S.; Vardar, E. Molecular Classification of Breast Carcinoma: From Traditional, Old-Fashioned Way to A New Age, and A New Way. J. Breast Health 2015, 11, 59–66. [Google Scholar] [CrossRef]
- Perou, C.M.; Sørile, T.; Eisen, M.B.; Van De Rijn, M.; Jeffrey, S.S.; Ress, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular Portraits of Human Breast Tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef]
- Sinn, H.P.; Kreipe, H. A Brief Overview of the WHO Classification of Breast Tumors, 4th Edition, Focusing on Issues and Updates from the 3rd Edition. Breast Care 2013, 8, 149–154. [Google Scholar] [CrossRef]
- Giuliano, A.E.; Connolly, J.L.; Edge, S.B.; Mittendorf, E.A.; Rugo, H.S.; Solin, L.J.; Weaver, D.L.; Winchester, D.J.; Hortobagyi, G.N. Breast Cancer-Major Changes in the American Joint Committee on Cancer Eighth Edition Cancer Staging Manual. CA Cancer J. Clin. 2017, 67, 290–303. [Google Scholar] [CrossRef]
- Fisusi, F.A.; Akala, E.O. Drug Combinations in Breast Cancer Therapy. Pharm. Nanotechnol. 2019, 7, 3–23. [Google Scholar] [CrossRef]
- Burstein, H.J.; Curigliano, G.; Thürlimann, B.; Weber, W.P.; Poortmans, P.; Regan, M.M.; Senn, H.J.; Winer, E.P.; Gnant, M.; Aebi, S.; et al. Customizing Local and Systemic Therapies for Women with Early Breast Cancer: The St. Gallen International Consensus Guidelines for Treatment of Early Breast Cancer 2021. Ann. Oncol. 2021, 32, 1216–1235. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Dai, A.; Tran, R.; Wang, J. Identifying MiRNA Biomarkers for Breast Cancer and Ovarian Cancer: A Text Mining Perspective. Breast Cancer Res. Treat. 2023, 201, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, H.; Gao, F. Identification of MiRNA Biomarkers for Breast Cancer by Combining Ensemble Regularized Multinomial Logistic Regression and Cox Regression. BMC Bioinform. 2022, 23, 434. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.G.; Davies, M.; Lowery, A.J.; Miller, N.; Kerin, M.J. The Role of MicroRNA as Clinical Biomarkers for Breast Cancer Surgery and Treatment. Int. J. Mol. Sci. 2021, 22, 8290. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Kim, Y.S.; Kang, K.N.; Kim, K.H.; Park, Y.J.; Kim, C.W. Multiple MicroRNAs as Biomarkers for Early Breast Cancer Diagnosis. Mol. Clin. Oncol. 2021, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Palacios, A.; Rojas Carvajal, A.M.; Núñez-Negrillo, A.M.; Cortés-Martín, J.; Sánchez-García, J.C.; Aguilar-Cordero, M.J. MicroRNA Dysregulation in Early Breast Cancer Diagnosis: A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2023, 24, 8270. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, Z.; Talkhabi, M.; Taleahmad, S. Identification of Potential MicroRNA Diagnostic Panels and Uncovering Regulatory Mechanisms in Breast Cancer Pathogenesis. Sci. Rep. 2022, 12, 20135. [Google Scholar] [CrossRef]
- Khadka, V.S.; Nasu, M.; Deng, Y.; Jijiwa, M. Circulating MicroRNA Biomarker for Detecting Breast Cancer in High-Risk Benign Breast Tumors. Int. J. Mol. Sci. 2023, 24, 7553. [Google Scholar] [CrossRef]
- Nguyen, T.H.N.; Nguyen, T.T.N.; Nguyen, T.T.M.; Nguyen, L.H.M.; Huynh, L.H.; Phan, H.N.; Nguyen, H.T. Panels of Circulating MicroRNAs as Potential Diagnostic Biomarkers for Breast Cancer: A Systematic Review and Meta-Analysis. Breast Cancer Res. Treat. 2022, 196, 1–15. [Google Scholar] [CrossRef]
- Huynh, K.Q.; Le, A.T.; Phan, T.T.; Ho, T.T.; Pho, S.P.; Nguyen, H.T.; Le, B.T.; Nguyen, T.T.; Nguyen, S.T. The Diagnostic Power of Circulating MiR-1246 in Screening Cancer: An Updated Meta-Analysis. Oxid. Med. Cell. Longev. 2023, 2023, 8379231. [Google Scholar] [CrossRef]
- Tiberio, P.; Gaudio, M.; Belloni, S.; Pindilli, S.; Benvenuti, C.; Jacobs, F.; Saltalamacchia, G.; Zambelli, A.; Santoro, A.; De Sanctis, R. Unlocking the Potential of Circulating MiRNAs in the Breast Cancer Neoadjuvant Setting: A Systematic Review and Meta-Analysis. Cancers 2023, 15, 3424. [Google Scholar] [CrossRef]
- Naeli, P.; Winter, T.; Hackett, A.P.; Alboushi, L.; Jafarnejad, S.M. The Intricate Balance between MicroRNA-Induced MRNA Decay and Translational Repression. FEBS J. 2023, 290, 2508–2524. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V.; Bartel, B.; Bartel, D.P.; Burge, C.B.; Carrington, J.C.; Chen, X.; Dreyfuss, G.; Eddy, S.R.; Griffiths-Jones, S.; Marshall, M.; et al. A Uniform System for MicroRNA Annotation. RNA 2003, 9, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. MiRBase: MicroRNA Sequences, Targets and Gene Nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef] [PubMed]
- Dziechciowska, I.; Dąbrowska, M.; Mizielska, A.; Pyra, N.; Lisiak, N.; Kopczyński, P.; Jankowska-Wajda, M.; Rubiś, B. MiRNA Expression Profiling in Human Breast Cancer Diagnostics and Therapy. Curr. Issues Mol. Biol. 2023, 45, 9500–9525. [Google Scholar] [CrossRef] [PubMed]
- Loh, H.Y.; Norman, B.P.; Lai, K.S.; Rahman, N.M.A.N.A.; Alitheen, N.B.M.; Osman, M.A. The Regulatory Role of MicroRNAs in Breast Cancer. Int. J. Mol. Sci. 2019, 20, 4940. [Google Scholar] [CrossRef] [PubMed]
- Najjary, S.; Mohammadzadeh, R.; Mokhtarzadeh, A.; Mohammadi, A.; Kojabad, A.B.; Baradaran, B. Role of MiR-21 as an Authentic Oncogene in Mediating Drug Resistance in Breast Cancer. Gene 2020, 738, 144453. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.X.; Lu, B.B.; Wang, H.; Cheng, Z.X.; Yin, Y.M. MicroRNA-21 Modulates Chemosensitivity of Breast Cancer Cells to Doxorubicin by Targeting PTEN. Arch. Med. Res. 2011, 42, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Tan, Z.; Hu, H.; Liu, H.; Wu, T.; Zheng, C.; Wang, X.; Luo, Z.; Wang, J.; Liu, S.; et al. MicroRNA-21 Promotes Breast Cancer Proliferation and Metastasis by Targeting LZTFL1. BMC Cancer 2019, 19, 738. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Ye, P.; Long, X. Differential Expression Profiles of the Transcriptome in Breast Cancer Cell Lines Revealed by Next Generation Sequencing. Cell. Physiol. Biochem. 2017, 44, 804–816. [Google Scholar] [CrossRef]
- Mohmmed, E.A.; Shousha, W.G.; EL-Saiid, A.S.; Ramadan, S.S. A Clinical Evaluation of Circulating MiR-106a and Raf-1 as Breast Cancer Diagnostic and Prognostic Markers. Asian Pac. J. Cancer Prev. 2021, 22, 3513–3520. [Google Scholar] [CrossRef]
- You, F.; Luan, H.; Sun, D.; Cui, T.; Ding, P.; Tang, H.; Sun, D. MiRNA-106a Promotes Breast Cancer Cell Proliferation, Clonogenicity, Migration, and Invasion Through Inhibiting Apoptosis and Chemosensitivity. DNA Cell Biol. 2019, 38, 198–207. [Google Scholar] [CrossRef]
- You, F.; Li, J.; Zhang, P.; Zhang, H.; Cao, X. MiR106a Promotes the Growth of Transplanted Breast Cancer and Decreases the Sensitivity of Transplanted Tumors to Cisplatin. Cancer Manag. Res. 2020, 12, 233–246. [Google Scholar] [CrossRef]
- Dinami, R.; Ercolani, C.; Petti, E.; Piazza, S.; Ciani, Y.; Sestito, R.; Sacconi, A.; Biagioni, F.; Le Sage, C.; Agami, R.; et al. MiR-155 Drives Telomere Fragility in Human Breast Cancer by Targeting TRF1. Cancer Res. 2014, 74, 4145–4156. [Google Scholar] [CrossRef]
- Roth, C.; Rack, B.; Müller, V.; Janni, W.; Pantel, K.; Schwarzenbach, H. Circulating MicroRNAs as Blood-Based Markers for Patients with Primary and Metastatic Breast Cancer. Breast Cancer Res. 2010, 12, R90. [Google Scholar] [CrossRef]
- Li, P.; Xu, T.; Zhou, X.; Liao, L.; Pang, G.; Luo, W.; Han, L.; Zhang, J.; Luo, X.; ** Breast Cancer Microenvironment Through Single-Cell Omics. Front. Immunol. 2022, 13, 868813. [Google Scholar] [CrossRef] [PubMed]
- Boo, L.; Ho, W.Y.; Ali, N.M.; Yeap, S.K.; Ky, H.; Chan, K.G.; Yin, W.F.; Satharasinghe, D.A.; Liew, W.C.; Tan, S.W.; et al. Phenotypic and MicroRNA Transcriptomic Profiling of the MDA-MB-231 Spheroid-Enriched CSCs with Comparison of MCF-7 MicroRNA Profiling Dataset. PeerJ 2017, 5, e3551. [Google Scholar] [CrossRef] [PubMed]
- Ahram, M.; Mustafa, E.; Zaza, R.; Abu Hammad, S.; Alhudhud, M.; Bawadi, R.; Zihlif, M. Differential Expression and Androgen Regulation of MicroRNAs and Metalloprotease 13 in Breast Cancer Cells. Cell Biol. Int. 2017, 41, 1345–1355. [Google Scholar] [CrossRef]
- Search for: Breast Cancer, Other Terms: Micro RNA, Micro RNA|Card Results|ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/search?cond=Breast%20Cancer&term=Micro%20RNA&intr=micro%20RNA (accessed on 2 November 2023).
- Lima, J.F.; Cerqueira, L.; Figueiredo, C.; Oliveira, C.; Azevedo, N.F. Anti-MiRNA Oligonucleotides: A Comprehensive Guide for Design. RNA Biol. 2018, 15, 338–352. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Function(s) | Estimated Risk | BC Type | Refs |
|---|---|---|---|---|
| BRCA1 | DNA repair transcription regulation cell cycle regulation chromatin remodeling | 55–65% by age 70 | TNBC luminal B | [11,25,27,28] |
| BRCA2 | DNA repair DNA replication transcription regulation cell cycle regulation mitophagy | ~45% by age 70 | TNBC luminal B | [11,25,28] |
| PALB2 | DNA repair | All women: RR 2.3, 95% CI 1.4–3.9 < 50 years: RR 3.0, 95% CI 1.4–5.5 | n/a | [17,25,28] |
| PTEN | cell survival cell growth | 85% lifetime | luminal A luminal B | [12,25,26,27,28] |
| TP53 | cell cycle regulation | 25% by age 74 | all | [13,25,26,27,28] |
| CDH1 | cell adhesion | 39% lifetime | luminal A | [14,25,26,27,28] |
| STK11 | cell cycle regulation | 32% by age 60 | n/a | [15,25,28] |
| CHEK2 | DNA repair cell cycle regulation apoptosis | Female: RR 1.70, 95% CI 1.3–2.2 Male: RR 10.3, 95% CI 3.5–30.0 | n/a | [16,25,26,27,28] |
| BRIP1 | DNA repair | All women: RR 2.0, 95% | n/a | [23,25] |
| ATM | DNA repair | RR 2.37, 95% CI 1.5–3.8 | n/a | [18,25,26,27,28] |
| Tumor | Node | Metastasis | |||
|---|---|---|---|---|---|
| Tx | no primary tumor information | Nx | not assessable | Mx | not assessed |
| T0 | no primary tumor evidence | N0 | no clinically positive nodes | M0 | no evidence |
| TIS | carcinoma in situ (primary sites) | N1 | single, ipsilateral, size < 3 cm | M1 | metastasis present at distance |
| T1 | size < 2 cm | N2a | single, ipsilateral, size 3–6 cm | ||
| T2 | size 2 to 4 cm | N2b | multiple, ipsilateral, size < 6 cm | ||
| T3 | size > 4 cm | N3 | massive/ipsilateral/bilateral/controlateral | ||
| T4 | size > 4 cm, pterygoid muscle, base of tongue or skin involved | N3a | ipsilateral node(s), one more than 6 cm | ||
| N3b | bilateral | ||||
| N4 | controlateral | ||||
| miR Name | Target Gene(s) | Affected Cellular Functions | Refs |
|---|---|---|---|
| miR-21 | PTEN | drug resistance | [65,66] |
| miR-21 | LZTFL1 | proliferation and metastasis | [67] |
| miR-21 | IGFBP3 TPM1 PCD4 TGF-β1 | proliferation, metastasis, epithelial-to-mesenchymal transition (EMT), apoptosis | [68] |
| miR-106a | RAF-1 | invasion and proliferation | [69] |
| miR-106a | P53 BAX RUNX3 Bcl-2 ABCG2 | proliferation, colony-forming capacity, migration, invasion, apoptosis, sensitivity to cisplatin | [70,71] |
| miR-155 | TRF1 | telomere fragility | [72] |
| miR-141 | ANP32E | migration and invasion | [74] |
| let-7 | ERCC6 | proliferation, apoptosis | [78] |
| miR-335 | ERα IGF1R SP1 ID4 | proliferation, apoptosis | [80] |
| miR-335 | c-Met | cell scattering, migration, and invasion | [81] |
| miR-126 | VEGFA PIK3R2 | angiogenesis, tumor genesis and growth | [83] |
| miR-126 | PIK3R2 | trastuzumab resistance | [84] |
| miR-199a/b-3p | PAK4 | migration and invasion | [86] |
| miR-199a-3p | mTOR c-Met | cell cycle progression, doxorubicin sensitivity, apoptosis | [87] |
| miR-199a-3p | TFAM | resistance to cisplatin | [88] |
| miR-199a-3p | TFAM | angiogenesis and metastasis under hypoxia | [89] |
| miR-101 | POMP Stmn1 DNMT3A EYA1 VHL SOX2 Jak2 MCL-1 | proliferation, apoptosis, angiogenesis, drug resistance, invasion, metastasis | [91] |
| miR-101-3p | COX-2 | migration, metastasis | [92] |
| miR-101-3p | EZH2 | migration, invasion, proliferation | [93] |
| miR-101-5p | GINS1 | DNA replication | [94] |
| miR-9 | FOXO1 | proliferation, migration, invasion | [99] |
| miR-9 | STARD13 | EMT, metastasis | [100] |
| miR-9 | LIFR | metastasis | [101] |
| miR-9 | elf5A2 | resistance to doxorubicin | [102] |
| miR-9 | HMGA2 EGR1 IGFBP3 | proliferation, metastasis, EMT, apoptosis | [68] |
| miR-9 | PDGFRβ | vasculogenesis | [103] |
| miR-200 | PDGFRβ | vasculogenesis | [103] |
| let-7a-5p miR-9-5p miR-10b miR-21 miR-22-3p miR-23b-3p miR-25-3p miR-29 miR-34a miR-93-5p miR-99a-5p/-3p miR-100-5p miR-101-3p miR-101-5p miR-126-5p/-3p miR-141-3p miR-143-5p/-3p miR-144-5p/-3p miR-145 miR-155 mir-181b1-5p miR-195-5p miR-199a-5p miR-200a miR-203 miR-203a-3p miR-205 miR-210-3p miR-221/222 miR-373 | n/a | biomarkers | [73,76,87,90,94,95,97,98] |
| miR | Organ | Target(s) | Notes | Refs |
|---|---|---|---|---|
| 125 | CNS | n/a | deregulated, pediatric | [130] |
| 125 | CNS | n/a | deregulated | [131,132] |
| 125 | CNS | p53, p38MAPK | none | [133] |
| 125 | CNS | BMF | none | [134] |
| 125a | ovary | n/a | EMT negative regulator | [144] |
| 125b | ovary | BCL3 | none | [145] |
| 125b | ovary | n/a | serum biomarker | [146] |
| 125b | bladder | E2F3 | none | [147] |
| 125b | bladder | n/a | urine biomarker | [148] |
| 125-3p | bladder | n/a | hypoxia regulated | [149] |
| 125 | bladder | n/a | survival predictor | [150] |
| 125a | liver | MMP11, VEGF | none | [151] |
| 125b | liver | Mcl-1, IL6R | none | [152] |
| 125b | liver | Lin28B2 | none | [153] |
| 125 | liver | Pokemon | none | [154] |
| 125 | liver | TRAF6 | none | [155] |
| 125 | liver | hexokinase II | none | [156] |
| 125 | liver | FOXM1 | none | [157] |
| 125 | skin | NCAM | none | [158] |
| 125 | skin | c-Jun | none | [159] |
| 125b | skin | MMP13 | none | [160] |
| 125b | skin | STAT3 | none | [161] |
| 125 | skin | n/a | deregulated | [162] |
| 125b | bone | STAT3 | none | [163,164] |
| 125 | bone | ErbB2 | none | [165] |
| 125 | bone | BAP1 | none | [166] |
| 125 | lung | n/a | survival predictor | [167] |
| 125 | lung | EGFR | none | [168] |
| 125 | lung | HER2 | trastuzumab resistance | [169] |
| 125 | lung | MMP13 | none | [170] |
| 125 | pancreas | n/a | deregulated | [171,172] |
| 125 | pancreas | NEDD9 | none | [173] |
| 125 | prostate | n/a | deregulated | [174,175,176] |
| 125 | prostate | BAK1 | none | [177] |
| 125 | prostate | p53, PUMA | none | [178] |
| 125b | thyroid | Foxp3 | cisplatin sensitivity | [179] |
| 125b | stomach | PPP1CA-Rb | none | [180] |
| 125a-5p | colon | BCL2, BCL2L12, MCL1 | none | [181] |
| 125b | kidney | n/a | survival predictor | [182] |
| miR | Reg. | Target | Reg. | Cellular Function | Cell Line | Ref. |
|---|---|---|---|---|---|---|
| miR-125a miR-125b | ↑ ↑ | ERBB2 ERBB3 | ↓ ↓ | migration invasion | SKBR3 | [189] |
| miR-125b | ↓ | ETS1 | ↑ | proliferation | BC samples | [190] |
| miR-125b | ↓ | MUC1 | ↑ | apoptosis | BT-549 ZR-75-1 | [191] |
| miR-125b | ↓ | STARD13 | ↑ | metastasis | MCF-7 MDA-MB-231 | [192] |
| miR-125 | ↓ | n/a | n/a | radioresistance | MCF-7 MDA-MB-231 | [193] |
| miR-125b | ↑ | n/a | n/a | chemoresistance proliferation apoptosis | blood samples | [194] |
| miR-125b | ↑ | BAK1 | ↓ | chemoresistance apoptosis | MDA-435 MDA-436 MDA-231 MCF7 SKBR3 | [195] |
| miR-125a-5p miR-125b | ↓ ↓ | n/a | n/a | age-dependent BC formation | BC samples | [196] |
| miR-125b | ↓ | MMP11 | ↑ | proliferation migration invasion | T47D SKBR3 | [197] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piergentili, R.; Marinelli, E.; Cucinella, G.; Lopez, A.; Napoletano, G.; Gullo, G.; Zaami, S. miR-125 in Breast Cancer Etiopathogenesis: An Emerging Role as a Biomarker in Differential Diagnosis, Regenerative Medicine, and the Challenges of Personalized Medicine. Non-Coding RNA 2024, 10, 16. https://doi.org/10.3390/ncrna10020016
Piergentili R, Marinelli E, Cucinella G, Lopez A, Napoletano G, Gullo G, Zaami S. miR-125 in Breast Cancer Etiopathogenesis: An Emerging Role as a Biomarker in Differential Diagnosis, Regenerative Medicine, and the Challenges of Personalized Medicine. Non-Coding RNA. 2024; 10(2):16. https://doi.org/10.3390/ncrna10020016
Chicago/Turabian StylePiergentili, Roberto, Enrico Marinelli, Gaspare Cucinella, Alessandra Lopez, Gabriele Napoletano, Giuseppe Gullo, and Simona Zaami. 2024. "miR-125 in Breast Cancer Etiopathogenesis: An Emerging Role as a Biomarker in Differential Diagnosis, Regenerative Medicine, and the Challenges of Personalized Medicine" Non-Coding RNA 10, no. 2: 16. https://doi.org/10.3390/ncrna10020016