
What about Exotic Species? Significance of Remains of Strange and Alien Animals in the Baltic Sea Region, Focusing on the Period from the Viking Age to High Medieval Times (800–1300 CE)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
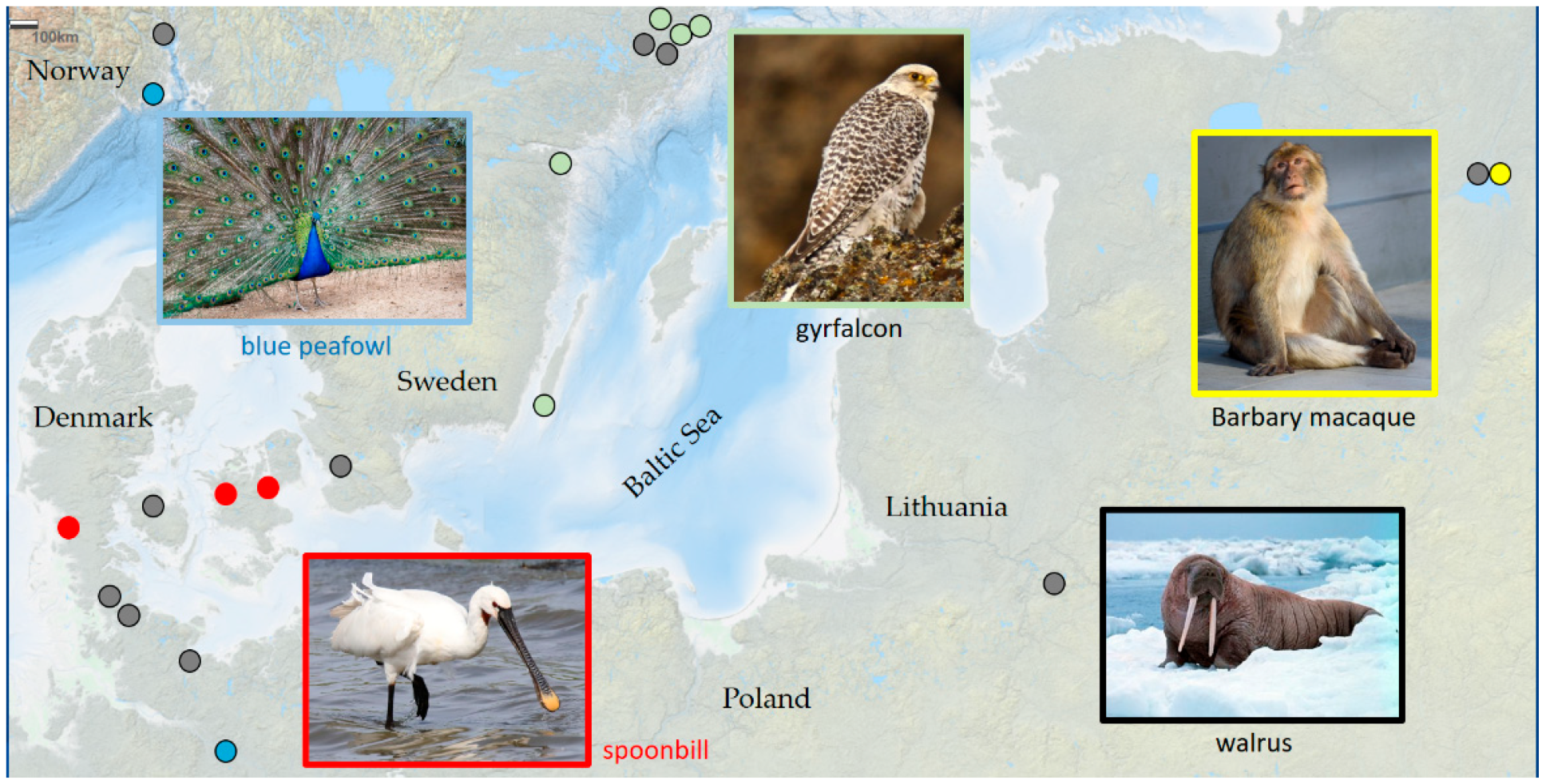
- They are the first of their kind in an area, in the course of natural expansion.
- They are the last of their kind in the area; the reasons for their disappearance are diverse.
- They appear as gifts or merchandise from distant regions.
- They existed only in the imagination of people, but their real existence was generally assumed.
2. Materials and Methods
3. Results
3.1. Natural or Anthropogenic Range Expansion: First of Their Kind
3.2. Natural or Anthropogenic Range Restriction: Last of Their Kind
3.3. Gifts or Merchandise from Distant Regions
3.4. Exotic Animals Existing Only in People’s Imagination
4. Discussion
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ashby, S.P. The role of zooarchaeology in the interpretation of socioeconomic status: A discussion with reference to medieval Europe. Archaeol. Rev. Cam. 2002, 18, 37–59. [Google Scholar]
- Van Neer, W.; Ervynck, A. Remains of traded fish in archaeological sites: Indicators of status, or bulk food? In Behaviour behind Bones: The Zooarchaeology of Ritual, Religion, Status, and Identity; O’Day, S.J., van Neer, S.W., Ervynck, A., Eds.; Oxbow Books: Oxford, UK, 2004; pp. 203–214. [Google Scholar]
- Schmölcke, U. Nutztierhaltung, Jagd und Fischfang. In Zur Nahrungsmittelwirtschaft des Frühgeschichtlichen Handelsplatzes von Groß Strömkendorf, Landkreis Nordwestmecklenburg; Archäologisches Landesmuseum Mecklenburg-Vorpommern: Lübstorf, Germany, 2004. [Google Scholar]
- Lightfoot, E.; Naum, M.; Kadakas, V.; Russow, E. The influence of social status and ethnicity on diet in mediaeval Tallinn as seen through stable isotope analysis. Est. J. Archaeol. 2016, 20, 81–107. [Google Scholar] [CrossRef] [Green Version]
- Błaszczyk, D.; Beaumont, J.; Krzyszowski, A.; Poliński, D.; Drozd-Lipińska, A.; Wrzesińska, A.; Wrzesiński, J. Social status and diet. Reconstruction of diet of individuals buried in some early medieval chamber graves from Poland by carbon and nitrogen stable isotopes analysis. J. Archaeol Sci. Rep. 2021, 38, 103103. [Google Scholar] [CrossRef]
- Oras, E.; Tõrv, M.; Jonuks, T.; Malve, M.; Radini, A.; Isaksson, S.; Gledhill, A.; Kekišev, O.; Vahur, S.; Leito, I. Social food here and hereafter: Multiproxy analysis of gender-specific food consumption in conversion period inhumation cemetery at Kukruse, NE-Estonia. J. Archaeol. Sci. 2018, 97, 90–101. [Google Scholar] [CrossRef]
- Twiss, K. The Archaeology of Food and Social Diversity. J. Archaeol. Res. 2012, 20, 357–395. [Google Scholar] [CrossRef]
- Heinrich, D.; von den Driesch, A.; Benecke, N. Holozängeschichte der Tierwelt Europas. Datacollection Ed; IANUS: Berlin, Germany, 2016. [Google Scholar] [CrossRef]
- Niethammer, G. Die Einbürgerung von Säugetieren und Vögeln in Europa. Ergebnisse und Aussichten; Paul Parey: Hamburg/Berlin, Germany, 1963. [Google Scholar]
- Nowak, E. Ausbreitung der Tiere; Ziemsen: Wittenberg Lutherstadt, Germany, 1977. [Google Scholar]
- Hengeveld, R. Dynamic Biogeography; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Cox, C.B.; Moore, P.D. Biogeography: An Ecological and Evolutionary Perspective, 5th ed.; Blackwell Scientific Publications: Oxford, UK, 1993. [Google Scholar]
- Kasparek, M. Dismigration und Brutarealexpansion der Türkentaube (Streptopelia decaocto). J. Ornithol. 1996, 137, 1–33. [Google Scholar] [CrossRef]
- Trouwborst, A.; Boitani, L.; Linnell, J.D. Interpreting ‘favourable conservation status’ for large carnivores in Europe: How many are needed and how many are wanted? Biodiv. Conserv. 2017, 26, 37–61. [Google Scholar] [CrossRef] [Green Version]
- Veedla, P. Rahvuslinnud ja Sümbollinnud Euroopas ja Kaugemal; Mõedaka: Pesapaik, Estonia, 2021. [Google Scholar]
- Dolata, P.T. The White Stork Ciconia ciconia protection in Poland by tradition, customs, law, and active efforts. In The White Stork in Poland: Studies in Biology, Ecology and Conservation; Tryjanowski, P., Sparks, T.H., Jerzak, L., Eds.; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2006; pp. 477–492. [Google Scholar]
- Gorobets, L.; Kovalchuk, O. Birds in the medieval culture and economy of the East Slavs in the 10–13th centuries AD. Environ. Archaeol. 2016, 22, 147–165. [Google Scholar] [CrossRef]
- Keller, V.; Herrando, S.; Voříšek, P.; Franch, M.; Kipson, M.; Milanesi, P.; Martí, D.; Anton, M.; Klvaňová, A.; Kalyakin, M.V.; et al. European Breeding Bird Atlas 2: Distribution, Abundance and Change. European Bird Census Council & Lynx Editions: Barcelona. 2020. Available online: https://ebba2.info/maps/species/Ciconia-ciconia/ebba2/breeding/ (accessed on 3 November 2022).
- von den Driesch, A.; Pöllath, N. Distribution history of European wild birds based on archaeozoological records. A contribution to human-bird relationships in the past. In Archaeobiodiversity. A European Perspective; Grupe, G., McGlynn, G., Peters, J., Eds.; Marie Leidorf: Rahden/Westf, Germany, 2011; pp. 71–216. [Google Scholar]
- Wigh, B. Animal Husbandry in the Viking Age Town of Birka and Its Hinterland. Ph.D. Thesis, Stockholm University, Stockholm, Sweden, 2001. [Google Scholar]
- Savinetsky, A.B.; Krylovich, O.A. On the history of the spread of the black rat (Rattus rattus L.; 1758) in northwestern Russia. Biol. Bull. 2011, 38, 203–207. [Google Scholar] [CrossRef]
- Hufthammer, A.K.; Walløe, L. Rats cannot have been intermediate hosts for Yersinia pestis during medieval plague epidemics in Northern Europe. J. Archaeol. Sci. 2013, 40, 1752–1759. [Google Scholar] [CrossRef]
- Tourunen, A. Animals in an Urban Context. A Zooarchaeological Study of the Medieval and Post-Medieval Town of Turku. Ph.D. Thesis, Turun University, Turku, Finland, 2008. [Google Scholar]
- Toplak, M. The Warrior and the Cat: A Re-Evaluation of the Roles of Domestic Cats in Viking Age Scandinavia. Curr. Swed. Archaeol. 2019, 27, 213–245. [Google Scholar] [CrossRef]
- Bitz-Thorsen, J.; Gotfredsen, A.B. Domestic cats (Felis catus) in Denmark have increased significantly in size since the Viking Age. Dan. J. Archaeol. 2018, 7, 241–254. [Google Scholar] [CrossRef]
- Skóra, K. Another Dimension of the Exotic? Notes on the Origins of the Domestic Cat in Central and Northern European Barbaricum during the Roman Period. In Rome and the Barbarians. An Interplay between Two Worlds; Olędzki, M., Dubicki, A., Eds.; Instytut Archeologii: Łódź, Poland, 2022; pp. 265–290. [Google Scholar]
- Engels, D.W. Classical Cats: The Rise and Fall of the Sacred Cat; Routledge: London; UK, 1999. [Google Scholar]
- Peters, J. Römische Tierhaltung und Tierzucht; Marie Leidorf: Rahden/Westfalen, Germany, 1998. [Google Scholar]
- Moilanen, U. Missä ovat Suomen rautakautiset kissat? Muinaistutkija 2020, 2020, 11–17. [Google Scholar]
- Petré, B. Arkeologiska Undersökningar på Fornlämning RAÄ 34, Lunda/Berga, Lovö sn.; Uppland Gravfält Från Vikingatid, Äldre Järnålder och Yngre Bronsålder Samt Boplatslämningar Från Bronsåldern; Lovö Archaeological Reports and Studies Nr 9 År 2010; Stockholm University: Stockholm, Sweden, 2010. [Google Scholar]
- Johansson, F.; Hüster, H. Untersuchungen an Skelettresten von Katzen aus Haithabu (Ausgrabung 1966–1996); Berichte über die Ausgrabungen in Haithabu 24; Karl Wachholtz Verlag: Neumünster, Germany, 1987. [Google Scholar]
- Łukaszewicz, K. The Ure-Ox. Ochr. Przyr. 1952, 20, 1–33. [Google Scholar]
- Aaris-Sørensen, K. The Holocene History of the Scandinavian Aurochs (Bos primigenius Bojanus, 1827). Wiss. Schr. Neanderthal. Mus. 1999, 1, 49–57. [Google Scholar]
- Rokosz, M. History of the Aurochs (Bos taurus primigenius) in Poland. Anim. Genet. Res. Inf. 1995, 16, 5–12. [Google Scholar] [CrossRef]
- Szafer, W. The Ure-ox, extinct in Europe since the seventeenth century: An early attempt at conservation that failed. Biol. Conserv. 1968, 1, 45–47. [Google Scholar] [CrossRef]
- Hufthammer, A.K. The Faunal Remains from the Gokstad Burial Mound Revised—Some New Results and Suggestions for Further Research. In Monumental Burials. The Art and Science of the Archaeological Revisit. The Gokstad Revitalised Project; Bill, J., Gjerde, H., Eds.; Aarhus University Press: Aarhus, Denmark, 2019; Volume 1. [Google Scholar]
- Gessner, J.; Spratte, S.; Kirschbaum, F. Historic Overview on the Status of the European Sturgeon (Acipenser sturio) and its Fishery in the North Sea and Its Tributaries with a Focus on German Waters. In Biology and Conservation of the European Sturgeon Acipenser sturio L.; Williot, P., Rochard, E., Desse-Berset, N., Kirschbaum, F., Gessner, J., Eds.; Springer: Heidelberg, Germany, 2011; pp. 195–219. [Google Scholar] [CrossRef]
- Nikulina, E.A.; Schmölcke, U. Reconstruction of the historical distribution of sturgeons (Acipenseridae) in the eastern North Atlantic based on ancient DNA and bone morphology of archaeological remains: Implications for conservation and restoration programmes. Divers. Distrib. 2016, 22, 1036–1044. [Google Scholar] [CrossRef]
- Nikulina, E.A.; Schmölcke, U. Historische Verbreitung von Europäischem Stör (Acipenser sturio) und Atlantischem Stör (Acipenser oxyrinchus) in West-und Mitteleuropa. Beitr. Archäozool. Prähist Anthr. 2018, 11, 37–44. [Google Scholar]
- Spratte, S. Störe in Schleswig-Holstein: Vergangenheit—Gegenwart—Zukunft; Landesamt für Landwirtschaft, Umwelt und Ländliche Räume des Landes Schleswig-Holstein (LLUR): Flintbek, Germany, 2015. [Google Scholar]
- Bengtson, S.-A. Breeding ecology and extinction of the Great Auk (**uinus impennis): Anecdotal evidence and conjectures. Auk 1984, 101, 1–12. [Google Scholar] [CrossRef]
- Thomas, J.E.; Carvalho, G.R.; Haile, J.; Rawlence, N.J.; Martin, M.D.; Ho, S.Y.; Sigfusson, A.; Josefsson, V.A.; Frederiksen, M.; Linnebjerg, J.F.; et al. Demographic reconstruction from ancient DNA supports rapid extinction of the great auk. eLife 2019, 8, e47509. [Google Scholar] [CrossRef] [PubMed]
- Hufthammer, A.K.; Hufthammer, K.O. The great auk in Norway: From common to locally extinct. Int. J. Osteoarchaeol. 2022, 1–10. [Google Scholar] [CrossRef]
- Risberg, L. Sveriges Fåglar. Aktuell Översikt över Deras Utbredning, Numerär och Flyttning Samt Något om Svensk Ornitologi; Sveriges Ornitologiska Förening: Stockholm, Sweden, 1990. [Google Scholar]
- Kjartansdóttir, K. The changing symbolic meaning of the extinct great auk and its afterlife as a museum object at the Natural History Museum of Denmark. J. Nord. Mus./Nord. Museol. 2019, 2, 41–56. [Google Scholar] [CrossRef]
- De Lozoya, A.V.; García, D.G.; Parish, J. A great auk for the Sun King. Arch. Nat. Hist. 2016, 43, 41–56. [Google Scholar] [CrossRef]
- Nikulina, E.A.; Schmölcke, U. First archaeogenetic results verify the mid-Holocene occurrence of Dalmatian pelican Pelecanus crispus far out of present range. J. Avian. Biol. 2015, 46, 344–351. [Google Scholar] [CrossRef]
- Gansum, T. The royal Viking Age ship grave from Gokstad in Vestfold, eastern Norway, and its link to falconry. In Raptor and Human—Falconry and Bird Symbolism Throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Advanced Studies on the Archaeology and History of Hunting; Wachholtz Verlag—Murmann Publishers: Kiel/Hamburg, Germany, 2018; Volumes 1.1–1.4, pp. 717–726. [Google Scholar]
- Cannell, R.J.S.; Bill, J.; Macphail, R. Constructing and deconstructing the Gokstad mound. Antiquity 2020, 94, 1278–1295. [Google Scholar] [CrossRef]
- Dobat, A.S. Viking stranger-kings: The foreign as a source of power in Viking Age Scandinavia, or, why there was a peacock in the Gokstad ship burial? Early Mediev. Eur. 2015, 23, 161–201. [Google Scholar] [CrossRef]
- Boessneck, J. Vogelknochenfunde aus der Burg auf dem Weinberg in Hitzacker/Elbe und dem Stadtkern von Dannenberg/Jeetzel (Mittelalter). Neue Ausgr. Niedersachs. 1982, 15, 345–394. [Google Scholar]
- Makowiecki, D.; Tomek, T.; Bochenski, Z.M. Birds in Early Medieval Greater Poland: Consumption and Hawking. Int. J. Osteoarch. 2014, 24, 358–364. [Google Scholar] [CrossRef]
- Makowiecki, D.; Zabilska-Kunek, M.; Seetah, K.; Jarzebowski, M.; Pluskowski, A. Farming, hunting and fishing in medieval Prussia: The Zooarchaeological data. In Terra Sacra: Environment, Colonisation and the Crusader States in Medieval Prussia and Livonia; Pluskowski, A., Ed.; Brepols Publishers: Turnhout, Belgium, 2019; pp. 333–376. [Google Scholar]
- Makowiecki, D.; Gotfredsen, A.B. Bird remains of Medieval and Post-Medieval coastal sites at the Southern Baltic Sea, Poland. Acta Zool. Crac. 2002, 45, 65–84. [Google Scholar]
- Gotfredsen, A.B. Birds in Subsistence and Culture at Viking Age Sites in Denmark. Int. J. Osteoarchaeol. 2014, 24, 365–377. [Google Scholar] [CrossRef]
- Grimm, O. From Aachen in the west to Birka in the north and Mikulčice in the east—Some archaeological remarks on bird of prey bones and falconry as being evidenced in premodern settlement contexts in parts of Europe (pre and post 1000 AD). In Raptor and Human—Falconry and Bird Symbolism Throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag—Murmann Publishers: Kiel/Hamburg, Germany, 2018; pp. 479–493. [Google Scholar]
- van Eerden, M.R. Patchwork. Patch Use, Habitat Exploitation and Carrying Capacity for Water Birds in Dutch Freshwater Wetlands. Ph.D. Thesis, Rijksuniversiteit Groningen, Groningen, The Netherlands, 1997. [Google Scholar]
- Orten Lie, R. Falconry, falcon-catching and the role of birds of prey in trade and as alliance gifts in Norway (800–1800 AD) with an emphasis on Norwegian and later foreign participants in falcon-catching. In Raptor and Human—Falconry and Bird Symbolism Throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag—Murmann Publishers: Kiel/Hamburg, Germany, 2018; pp. 727–786. [Google Scholar]
- Ægisson, S. Icelandic Trade with Gyrfalcons from Medieval Times to the Modern Era; Sigurður Ægisson: Siglufjörður, Iceland, 2015. [Google Scholar]
- Mehler, N.; Küchelmann, H.C.; Holterman, B. The export of gyrfalcons from Iceland during the 16th century: A boundless business in a proto-globalized world. In Raptor and Human—Falconry and Bird Symbolism Throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag—Murmann Publishers: Kiel/Hamburg, Germany, 2018; pp. 995–1020. [Google Scholar]
- Bochenski, Z.M.; Tomek, T.; Wertz, K.; Wojenka, M. Indirect evidence of falconry in medieval Poland as inferred from published zooarchaeological studies. Int. J. Osteoarchaeol. 2016, 26, 661–669. [Google Scholar] [CrossRef]
- Boessneck, J.; von den Driesch, A. Eketorp. Befestigung und Siedlung auf Öland/Schweden. Die Fauna; Royal Academy of Letters, History and Antiquities: Stockholm, Sweden, 1979. [Google Scholar]
- Ericson, P.G.; Tyrberg, T. The Early History of the Swedish Avifauna: A Review of the Subfossil Record and Early Written Sources; Almqvist & Wiksell International: Stockholm, Sweden, 2004. [Google Scholar]
- Vretemark, M. Birds of Prey as Evidence for Falconry in Swedish Burials and Settlements (550–1500 AD). In Raptor and Human—Falconry and Bird Symbolism Throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag—Murmann Publishers: Kiel/Hamburg, Germany, 2018; pp. 827–839. [Google Scholar]
- Brisbane, M.; Hambleton, E.; Maltby, M.; Nosov, E. A monkey’s tale: The skull of a macaque found at Ryurik Gorodishche during excavations in 2003. Mediev. Archaeol. 2007, 51, 185–190. [Google Scholar]
- Ahrland, Å. Vert and Venison—High Status Hunting and Parks in Medieval Sweden, in Hunting in Northern Europe until 1500 AD. Old Traditions and Regional Developments, Continental Sources and Continental Influences; Grimm, O., Schmölcke, U., Eds.; Wachholtz Verlag: Neumünster, Germany, 2013; pp. 439–464. [Google Scholar]
- Ahrland, Å. Power and paradise. Swedish deer parks in a long-term perspective. Bebygg. Tidskr. 2011, 61, 68–89. [Google Scholar]
- Andrén, A. Paradise lost: Looking for deer parks in medieval Denmark and Sweden. In Visions of the Past: Trends and Traditions in Swedish Medieval Archaeology; Andersson, P.H., Carelli, L.E., Eds.; Riksantikvarieämbetet: Stockholm, Sweden, 1997; pp. 469–490. [Google Scholar]
- Kocks, B. Die Tierknochenfunde aus den Burgen auf dem Weinberg in Hitzacker/Elbe und in Dannenberg (Mittelalter), I. Die Nichtwiederkäuer. Ph.D. Thesis, Ludwig Maximilians-Universität, München, Germany, 1978. [Google Scholar]
- Mulsow, R. Von der mittelalterlichen Universitas zur reformierten humanistischen Hochschule: Archäologische Funde des späten 16. Jahrhunderts aus der Blütezeit der Rostocker Universität. Mitt. Dtsch. Ges. Archäologie Mittelalt. Neuzeit 2007, 18, 59–70. [Google Scholar]
- Kahlow, S. Exotic Animals. Thoughts about Supply and Demand Based on Archaeological finds. In Transfer between Sea and Land. Maritime Vessels for Cultural Exchanges in the Early Modern Period; Kahlow, S., Ed.; Sidestone Press: Leiden, Belgium, 2018; pp. 87–118. [Google Scholar]
- Keighley, X.; Tange Olsen, M.; Jordan, P. Integrating cultural and biological perspectives on long-term human-walrus (Odobenus rosmarus rosmarus) interactions across the North Atlantic. Quatern. Res. 2019, 108, 5–25. [Google Scholar] [CrossRef]
- Frei, K.M.; Coutu, A.N.; Smiarowski, K.; Harrison, R.; Madsen, C.K.; Arneborg, J.; Frei, R.; Guðmundsson, G.; Sindbæk, S.M.; Woollett, J.; et al. Was it for walrus? Viking Age settlement and medieval walrus ivory trade in Iceland and Greenland. World Archaeol. 2015, 47, 439–466. [Google Scholar] [CrossRef]
- Barrett, J.H. The exploitation of walrus ivory in medieval Europe. In The Atlantic Walrus: Multidisciplinary Insights into Human–Animal Interactions; Keighley, X., Olsen, M.T., Jordan, P., Desjardins, S., Eds.; Academic Press: Amsterdam, The Netherlands, 2021; pp. 169–196. [Google Scholar]
- Barrett, J.H.; Boessenkool, S.; Kneale, C.J.; O’Connell, T.C.; Star, B. Ecological globalisation, serial depletion and the medieval trade of walrus rostra. Quat. Sci. Rev. 2020, 229, 20212773. [Google Scholar] [CrossRef]
- Brandt, L.Ø.; Haase, K.; Collins, M.J. Species identification using ZooMS, with reference to the exploitation of animal resources in the medieval town of Odense. Dan. J. Archaeol. 2018, 7, 139–153. [Google Scholar] [CrossRef]
- Keighley, X.; Palsson, S.; Einarsson, B.F.; Petersen, A.; Fernandez-Coll, M.; Jordan, P.; Olsen, M.T.; Malmquist, H.J. Disappearance of Icelandic Walruses Coincided with Norse Settlement. Mol. Biol. Evol. 2019, 36, 2656–2667. [Google Scholar] [CrossRef] [Green Version]
- Dectot, X. When ivory came from the seas. On some traits of the trade of raw and carved sea-mammal ivories in the Middle Ages. Anthropozoologica 2018, 53, 159–174. [Google Scholar] [CrossRef]
- Heizmann, W. Midgardschlange. In Mittelalter Mythen, 2. Dämonen, Monster, Fabelwesen; Müller, U., Wunderlich, W., Eds.; UVK Verlagsgesellschaft: St. Gallen, Switzerland, 1999; pp. 413–438. [Google Scholar]
- Pesch, A. Die Kraft der Tiere. Völkerwanderungszeitliche Goldhalskragen und Die Grundsätze Germanischer Kunst; Verlag des Römisch-Germanischen Zentralmuseums: Mainz, Germany, 2015. [Google Scholar]
- Pluskowski, A. Narwhals or Unicorns? Exotic Animals as Material Culture in Medieval Europe. Europ. J. Archaeol. 2004, 7, 291–313. [Google Scholar] [CrossRef]
- Bartosiewicz, L.; Choyke, A.M. Animals Step** off the Page. In Medieval Animals on the Move: Between Body and Mind; Bartosiewicz, L., Choyke, A.M., Eds.; Palgrave: Cham, Switzerland, 2020; pp. 1–12. [Google Scholar]
- Christen, A.G.; Christen, J.A. The unicorn and the narwhal: A tale of the tooth. J. Hist. Dent. 2011, 59, 135–142. [Google Scholar] [PubMed]
- Pansini, S.R. The Unicorn Tapestries: Religion, Mythology, and Sexuality in Late Medieval Europe. Ph.D. Thesis, Harvard University, Cambridge, UK, 2021. [Google Scholar]
- Katona, C. Exotic Encounters: Vikings and Faraway Species in Motion. In Medieval Animals on the Move; Bartosiewicz, L., Choyke, A.M., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 167–177. [Google Scholar]
- Balon, E.K. Origin and domestication of the wild carp, Cyprinus carpio: From Roman gourmets to the swimming flowers. Aquaculture 1995, 129, 3–48. [Google Scholar] [CrossRef]
- Hoffman, R.C. Environmental change and the culture of common carp in medieval Europe. Guelph Ichthyol. Rev. 1995, 3, 57–85. [Google Scholar]
- Ericson, P.G.P.; Tyrberg, T.; Kjellberg, A.S.; Jonsson, L.; Ullén, I. The earliest record of house sparrows (Passer domesticus) in northern Europe. J. Archaeol. Sci. 1997, 24, 183–190. [Google Scholar] [CrossRef]
- Reichstein, H.; Pieper, H. Untersuchungen zu Skelettresten von Vögeln aus Haithabu (Ausgrabung 1966–1969); Berichte über die Ausgrabungen in Haithabu 22; Karl Wachholtz Verlag: Neumünster, Germany, 1986. [Google Scholar]
- Corsi, M.R. Elite Networks and Courtly Culture in Medieval Denmark: Denmark in Europe, 1st to 14th Centuries. Ph.D. Thesis, University of Houston, Houston, TX, USA, 2014. [Google Scholar]
- Grane, T. Roman Imports in Scandinavia: Their Purpose and Meaning? In Rome beyond Its Frontiers. Imports, Attitudes, and Practices; Wells, P.S., Ed.; JRA: Portsmouth, UK, 2013; pp. 29–44. [Google Scholar]
- Aannestad, H.L. The allure of the foreign. The social and cultural dimension of imports in Scandinavia in the Viking Age. Viking Mediev. Scand. 2018, 14, 1–19. [Google Scholar] [CrossRef]
- Bomgardner, D.L. The trade in wild beasts for Roman spectacles: A green perspective. Anthropozoologica 1992, 16, 161–166. [Google Scholar]
- MacKinnon, M. Supplying Exotic Animals for the Roman Amphitheatre Games: New Reconstructions combining Archaeological, Ancient Textual, Historical and Ethnographic Data. Mouseion 2006, 6, 137–162. [Google Scholar] [CrossRef]
- Lindberg, N. The Emperor and His Animals: The Acquisition of Exotic Beasts for Imperial venationes. Greece Rome 2019, 66, 251–263. [Google Scholar] [CrossRef]
- Sommer, R.S.; Benecke, N.; Lõugas, L.; Nelle, O.; Schmölcke, U. Holocene survival of the wild horse in Europe: A matter of open landscape? J. Quatern. Sci. 2011, 26, 805–812. [Google Scholar] [CrossRef]
- Sommer, R.S.; Hegge, C.; Schmölcke, U. Lack of support for adaptation of post-glacial horses to woodlands. Nat. Ecol. Evol. 2018, 2, 582–583. [Google Scholar] [CrossRef]
- van Dam, P.J.E.M. New habitats for the rabbit in Northern Europe, 1300–1600. In Inventing Medieval Landscapes: Senses of Place in Western Europe; Howe, J., Wolfe, M., Eds.; University Press of Florida: Gainesville, FL, USA, 2002; pp. 57–69. [Google Scholar]
- Skyrienė, G.; Paulauskas, A. Distribution of invasive muskrats (Ondatra zibethicus) and impact on the ecosystem. Ekologija 2012, 58, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Kauhala, K.; Kowalczyk, R. Invasion of the raccoon dog Nyctereutes procyonoides in Europe: History of colonization, features behind its success, and threats to native fauna. Curr. Zool. 2011, 57, 584–598. [Google Scholar] [CrossRef] [Green Version]
- Muster, C.; Meyer, M.; Sattler, T. Spatial Arrangement Overrules Environmental Factors to Structure Native and Non-Native Assemblages of Synanthropic Harvestmen. PLoS ONE 2014, 9, e90474. [Google Scholar] [CrossRef] [Green Version]
- Hulme, P.E. Biological invasions in Europe: Drivers, pressures, states, impacts and responses. In Biodiversity under Threat; Hester, R.E., Harrison, R.M., Eds.; RSC Publishing: Cambridge, UK, 2007; pp. 56–80. [Google Scholar]
- Hulme, P.E.; Nentwig, W.; Pyšek, P.; Vilà, M. (Eds.) DAISIE, the Handbook of Alien Species in Europe; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Krajcarz, M.; Van Neer, W.; Krajcarz, M.T.; Popović, D.; Baca, M.; De Cupere, B.; Goffette, Q.; Küchelmann, H.C.; Gręzak, A.; Iwaszczuk, U.; et al. Stable isotopes unveil one millennium of domestic cat paleoecology in Europe. Sci. Rep. 2022, 12, 12775. [Google Scholar] [CrossRef]
- Keller, R.P.; Geist, J.; Jeschke, J.M.; Kühn, I. Invasive species in Europe: Ecology, status, and policy. Environ. Sci. Eur. 2011, 23, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. Camb. Philos. Soci. 2020, 95, 1511–1534. [Google Scholar] [CrossRef]
- Haubrock, P.J.; Turbelin, A.J.; Cuthbert, R.N.; Novoa, A.; Taylor, N.G.; Angulo, E.; Ballesteros-Mejia, L.; Bodey, T.W.; Capinha, C.; Diagne, C.; et al. Economic costs of invasive alien species across Europe. NeoBiota 2021, 67, 153–190. [Google Scholar] [CrossRef]
- Berger, J. Anthropogenic extinction of top carnivores and interspecific animal behaviour: Implications for the rapid decoupling of a web involving wolves, bears, moose and ravens. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1999, 266, 2261–2267. [Google Scholar] [CrossRef]
- Berger, J.; Swenson, J.E.; Persson, I.L. Recolonizing carnivores and naive prey: Conservation lessons from Pleistocene extinctions. Science 2001, 291, 1036–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, R.; Linnell, J.D.C.; Solberg, E.J. The future role of large carnivores in terrestrial trophic interactions: The north temperate view. In Large Herbivore Ecology, Ecosystem Dynamics and Conservation; Danell, K., Bergström, R., Duncan, P., Pastor, J., Eds.; Cambridge University Press: Cambridge, UK, 2006; pp. 413–448. [Google Scholar]
- Bocherens, H.; Hofman-Kaminska, E.; Drucker, D.G.; Schmölcke, U.; Kowalczyk, R. European Bison as a Refugee Species? Evidence from Isotopic Data on Early Holocene Bison and Other Large Herbivores in Northern Europe. PLoS ONE 2015, 10, e0115090. [Google Scholar] [CrossRef] [PubMed]
- Hofman-Kamińska, E.; Merceron, G.; Bocherens, H.; Makowiecki, D.; Piličiauskienė, G.; Ramdarshan, A.; Berlioz, E.; Kowalczyk, R. Foraging habitats and niche partitioning of European large herbivores during the Holocene—Insights from 3D dental microwear texture analysis. Palaeogeogr. Palaeoclim. Palaeoecol. 2018, 506, 183–195. [Google Scholar] [CrossRef]
- Cromsigt, J.P.G.M.; Kemp, Y.J.M.; Rodriguez, E.; Kivit, H. Rewilding Europe’s large grazer community: How functionally diverse are the diets of European bison, cattle, and horses? Restor. Ecol. 2018, 26, 891–899. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmölcke, U. What about Exotic Species? Significance of Remains of Strange and Alien Animals in the Baltic Sea Region, Focusing on the Period from the Viking Age to High Medieval Times (800–1300 CE). Heritage 2022, 5, 3864-3880. https://doi.org/10.3390/heritage5040199
Schmölcke U. What about Exotic Species? Significance of Remains of Strange and Alien Animals in the Baltic Sea Region, Focusing on the Period from the Viking Age to High Medieval Times (800–1300 CE). Heritage. 2022; 5(4):3864-3880. https://doi.org/10.3390/heritage5040199
Chicago/Turabian StyleSchmölcke, Ulrich. 2022. "What about Exotic Species? Significance of Remains of Strange and Alien Animals in the Baltic Sea Region, Focusing on the Period from the Viking Age to High Medieval Times (800–1300 CE)" Heritage 5, no. 4: 3864-3880. https://doi.org/10.3390/heritage5040199