3.1. Genes Implicated in DMT2 Disease According to DisGeNET Database
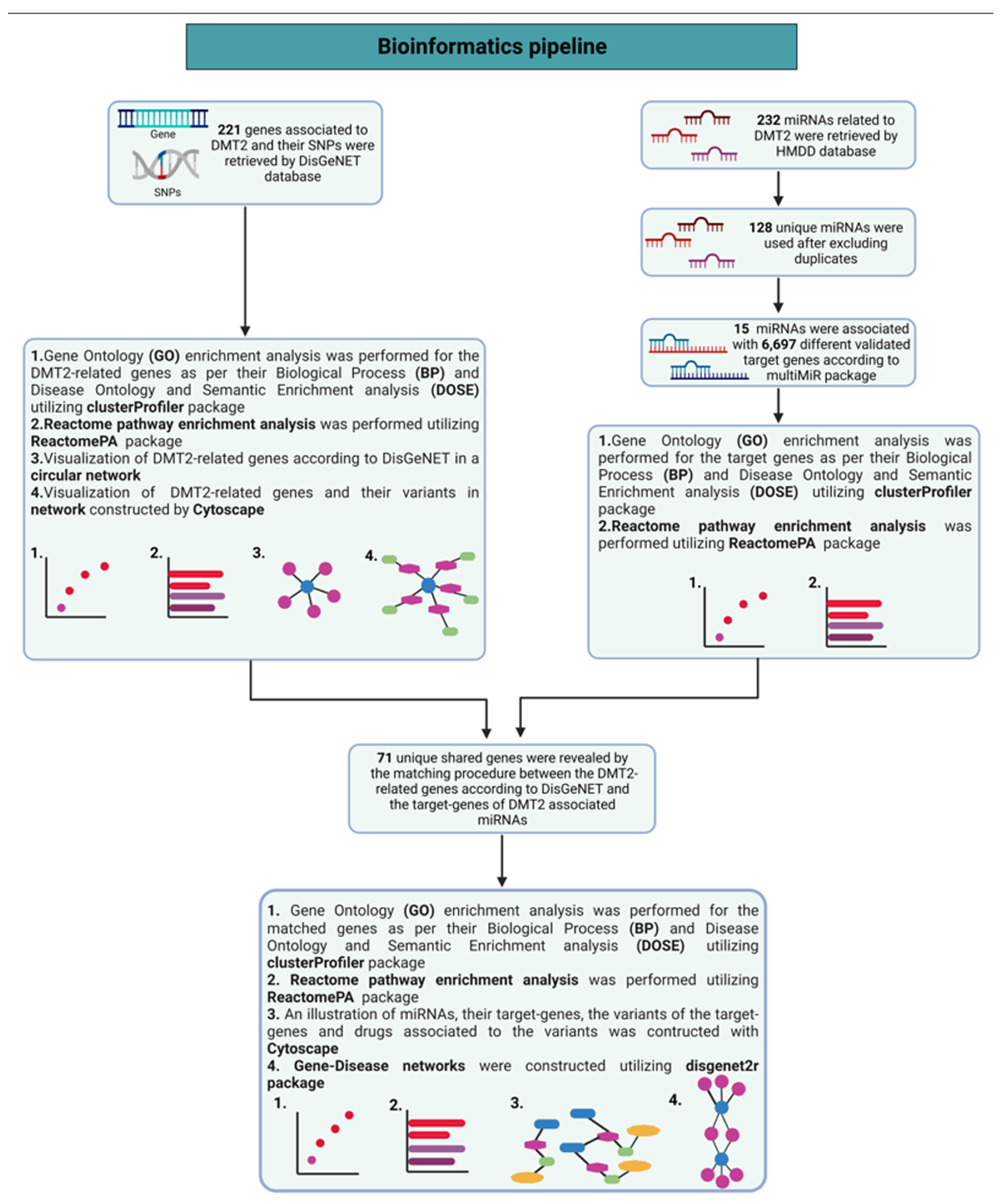
Our search in the DisGeNET database, using expert-curated data, revealed that 221 genes are related with DMT2 pathophysiological mechanisms (
Table S1 and
Figure 1a). In addition, the identified variants of the genes implicated in DMT2 are depicted in a circular network (
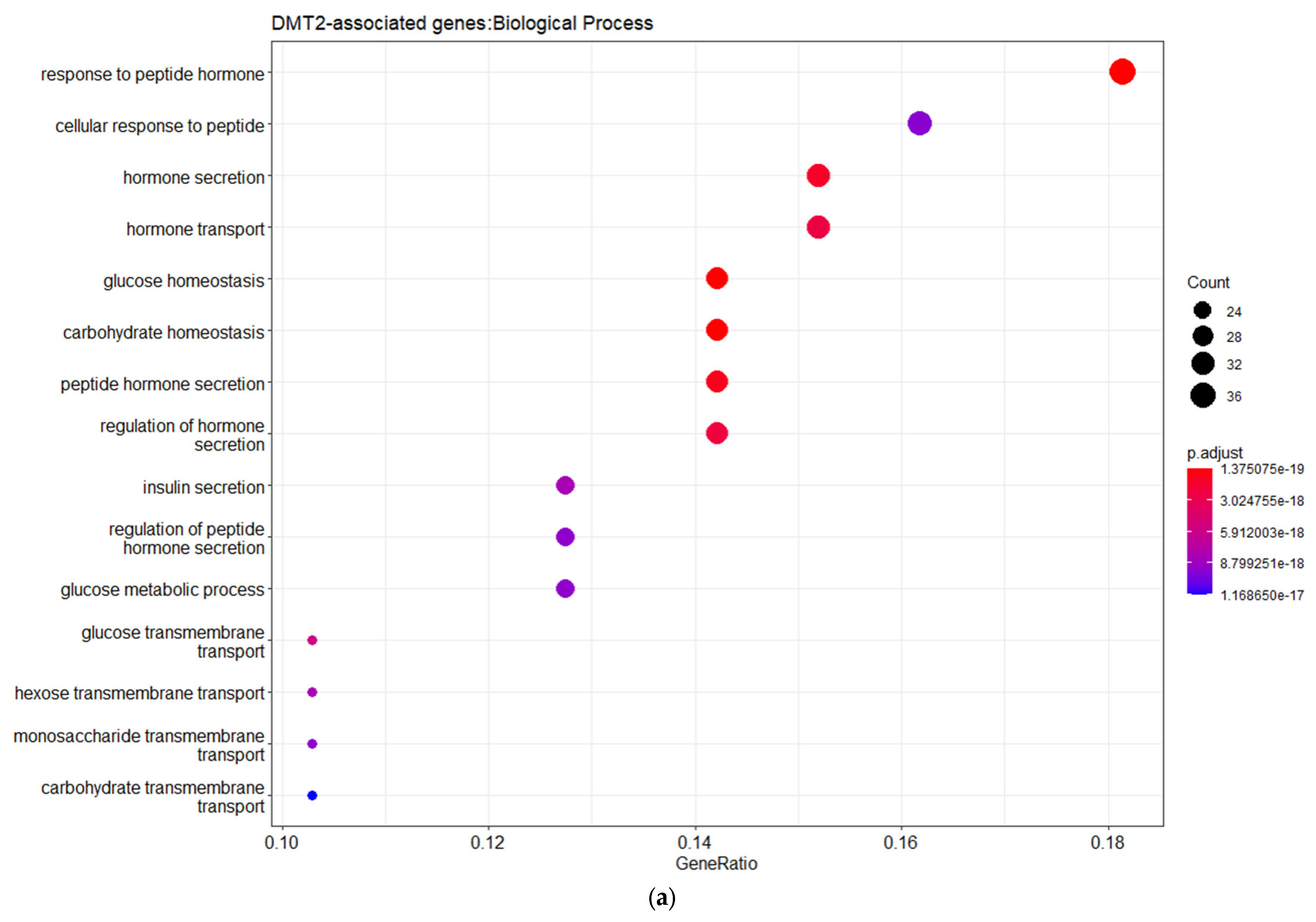
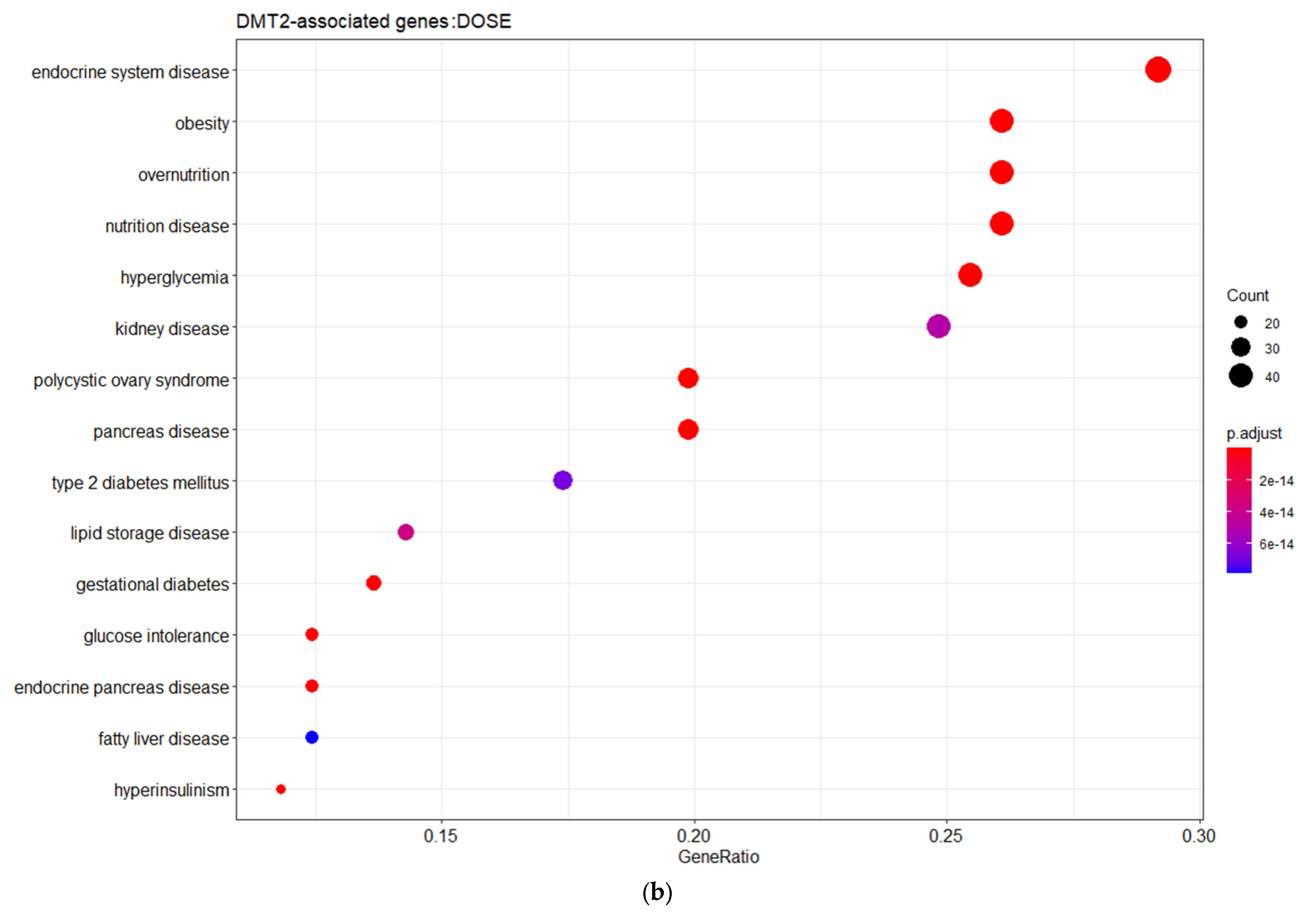
Figure 1b). The gene ontology (GO) enrichment analysis based on the biological process (BP) revealed that the DMT2-associated genes are implicated in cellular responses to peptide hormones, including insulin, as well as glucose homeostasis, transmembrane transport, and metabolic processes. The disease ontology (DOSE) enrichment analysis unveiled that the DMT2-related genes are implicated in diseases that usually co-exist in patients with DMT2, including glucose intolerance, obesity, kidney or pancreas diseases, fatty liver disease, and polycystic ovary syndrome.
Obesity is associated with an increased risk of develo** insulin resistance and is considered as a significant inducer of DMT2 evolution. In individuals with obesity, the adipose tissue releases increased levels of non-esterified fatty acids (NEFAs), hormones, glycerol, and pro-inflammatory cytokines that are implicated in insulin resistance. Obesity-associated insulin resistance and abnormalities in pancreatic islet-β cell function result in abnormal blood glycose levels and DMT2 development [
18]. In addition, insulin resistance in skeletal muscle is a dominant etiological factor of hyperlipidemia and excess fat accumulation in patients with DMT2 [
19]. The prevalence of chronic kidney disease (CKD) is two-to-five times higher in people with DMT2 as compared with non-diabetic individuals, since hyperglycemia promotes pathological changes withing the kidney [
20,
21]. Patients with nonalcoholic fatty liver disease (NAFLD) have an increased risk of DMT2 development and vice versa; NAFLD and DMT2 have similar pathophysiological pathways and insulin resistance is the hallmark presented in both diseases [
22]. The accumulation of toxic metabolites derived from triglycerides results in NAFLD-associated lipotoxicity. Such toxic metabolites accumulated in liver, pancreas, and muscles trigger inflammatory cascades and hepatic insulin resistance, which are pivotal inducers of DMT2 development [
23]. Non-alcoholic fatty pancreas disease (NAFPD) is also associated with obesity, metabolic syndrome (MetS), atherosclerosis, and DMT2, verifying the existence of molecular pathways that are common between the diseases [
24]. Impaired glycose homeostasis is also observed in patients with polycystic ovary syndrome (PCOS). This endocrine disorder usually coexists with prediabetes, which usually gradually develops into DMT2 over time. Insulin resistance and pancreatic β-cell dysfunction, which are the main pathophysiological factors for DMT2 development, are also concomitant in women with PCOS [
25].
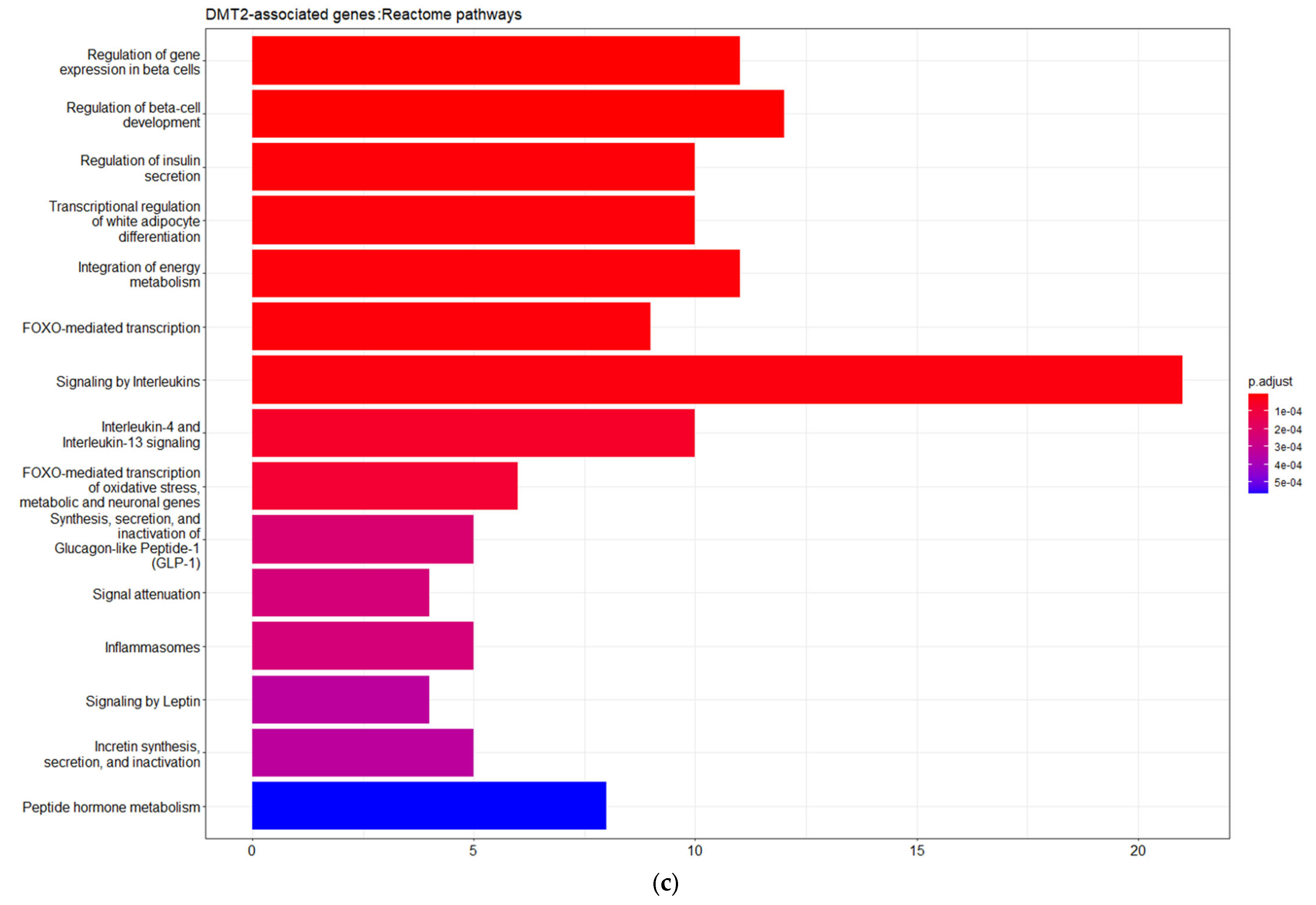
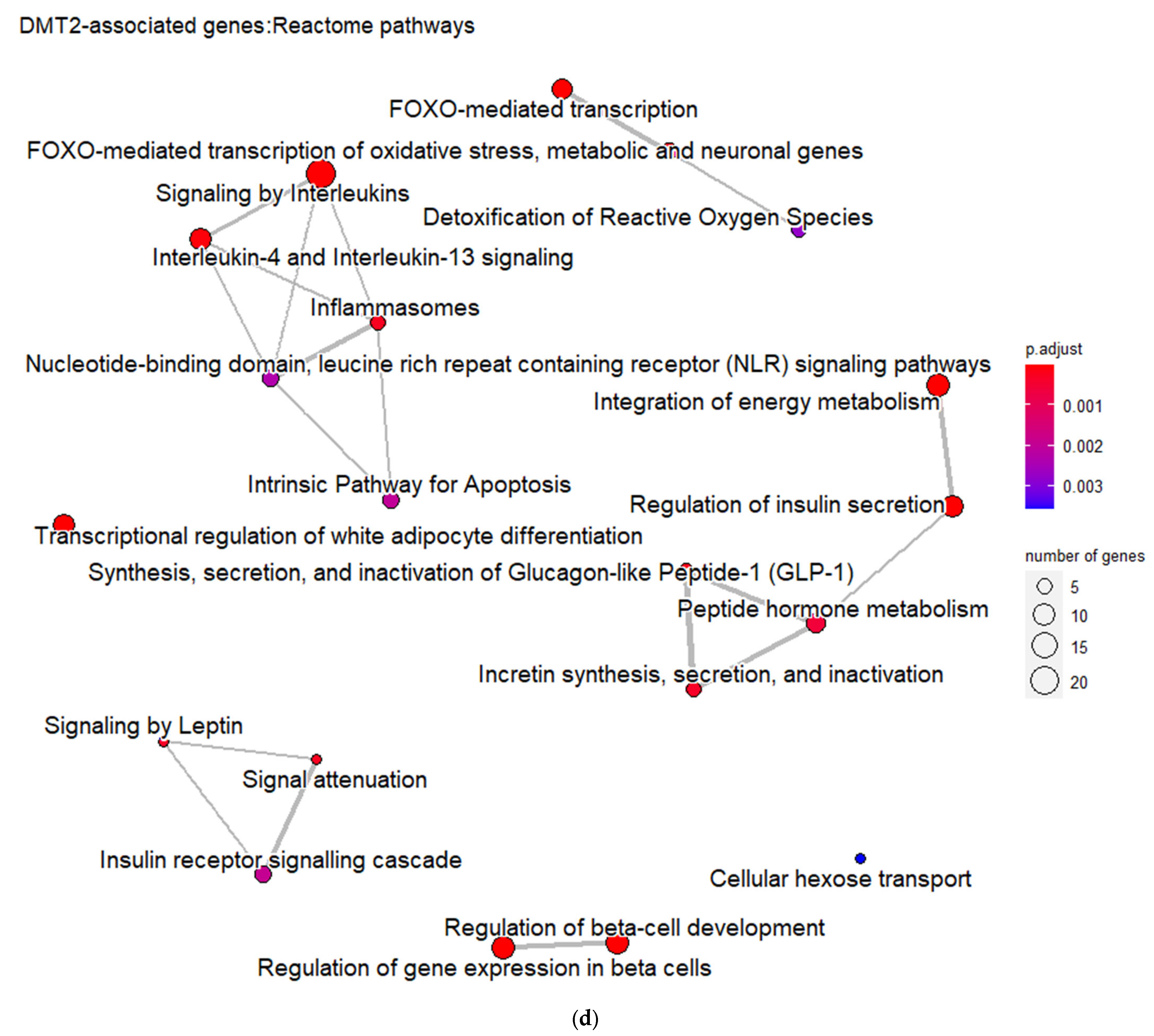
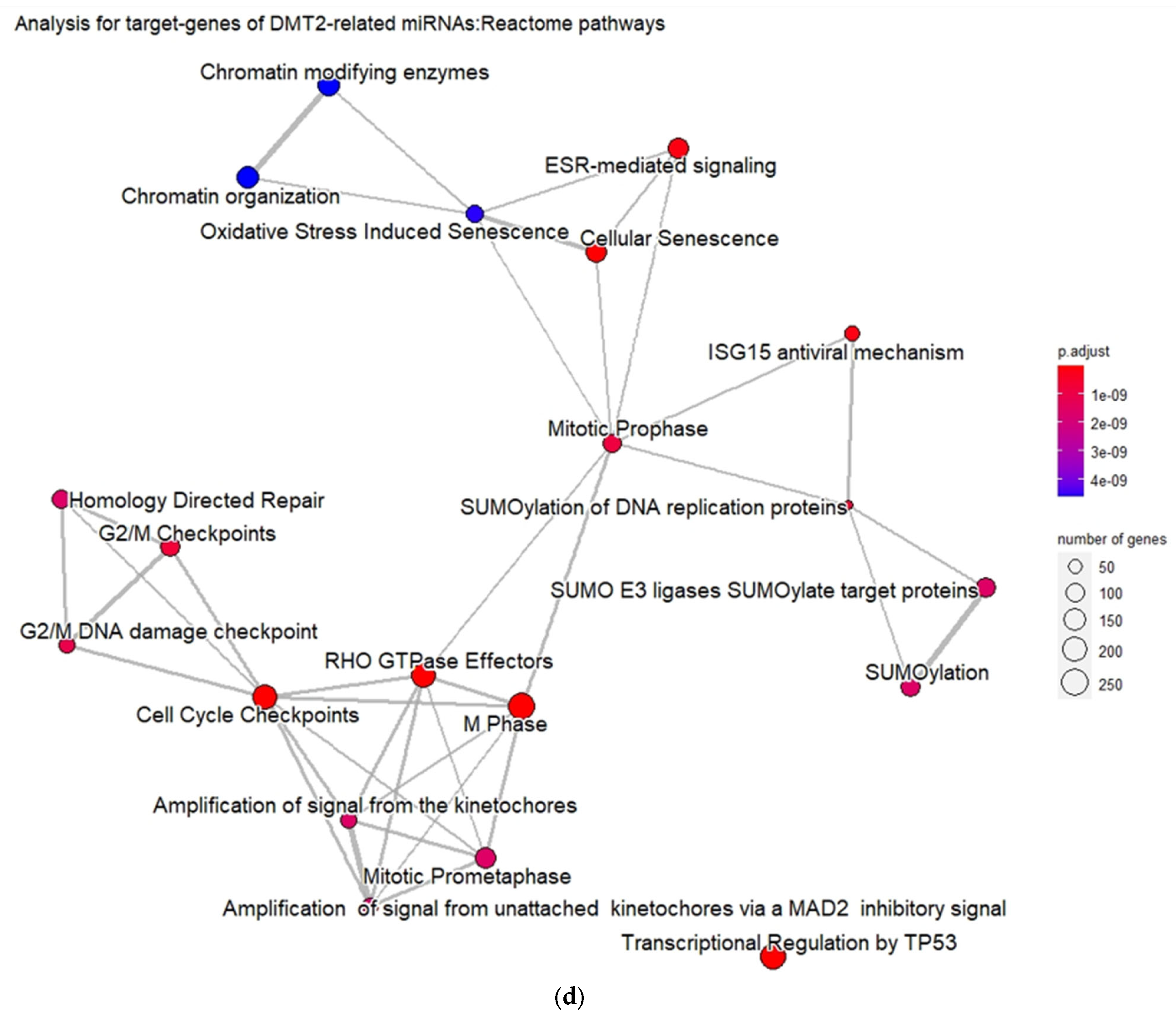
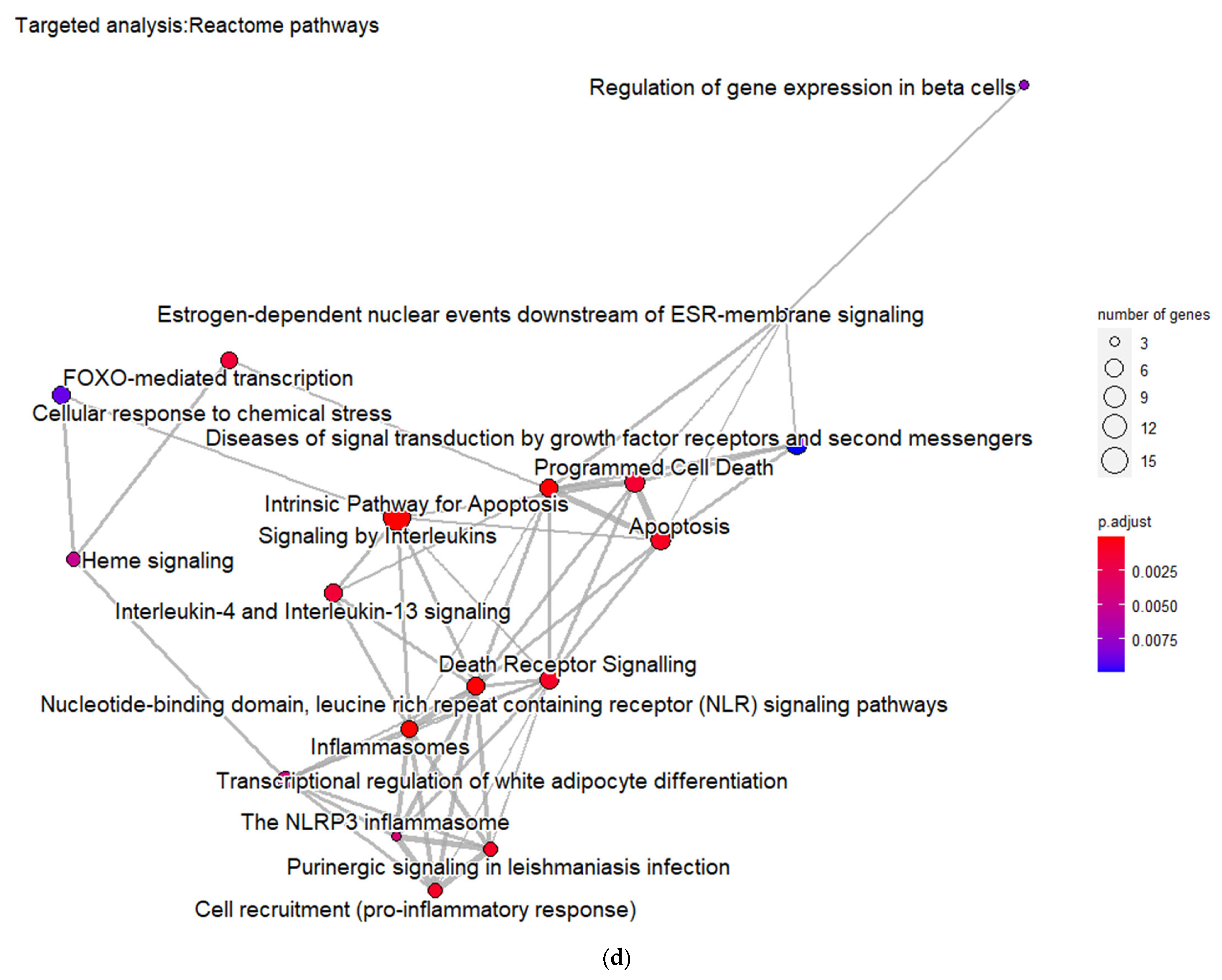
Furthermore, the DMT2-related genes are significantly enriched in pathways related to the regulation of gene expression in pancreatic β-cells, insulin secretion, metabolic hormones regulation, and interleukin signaling. These pathways form six different, larger pathway clusters, i.e., as depicted on
Figure 2d, mainly referring to: (a) Oxidative stress; (b) Interleukin-signaling inflammasomes and adipocyte differentiation; (c) Energy and hormone metabolism; (d) Leptin and insulin signaling; (e) β-pancreatic cell development and gene regulation; and (f) Cellular hexose transport. The abnormal function of the genes implicated in these pathway clusters is the driving etiological factor towards DMT2 development. Oxidative stress and chronic inflammation participate in the development of DMT2. Inflammatory mediators, including the interleukins 1-beta and 6 and tumor necrosis factor-alpha, in combination with hyperglycemia, induce the generation of reactive oxygen species implicated in DMT2 pathogenesis [
26]. Abnormalities in signaling pathways involved in energy and hormone metabolism, leptin and insulin signaling, and cellular glucose transport strongly determine the molecular signature associated with DMT2 development and progression [
27].
3.2. Experimentally Validated Target Genes of DMT2-Related miRNAs
The search of the HMDD database revealed 232 total miRNA entries that are related to DMT2. After excluding duplicates, we identified 128 unique miRNAs to be associated with DMT2. Amongst them, 16 miRNAs are already shown to be associated with their target genes, according to the multiMiR package. Subsequently, the miRNA-target gene interactions, which are not supported by experimental validation, were excluded. After applying this filtering, 15 miRNAs interacting with 6697 different target genes, according to available experimental validation data, were used for downstream analysis (see
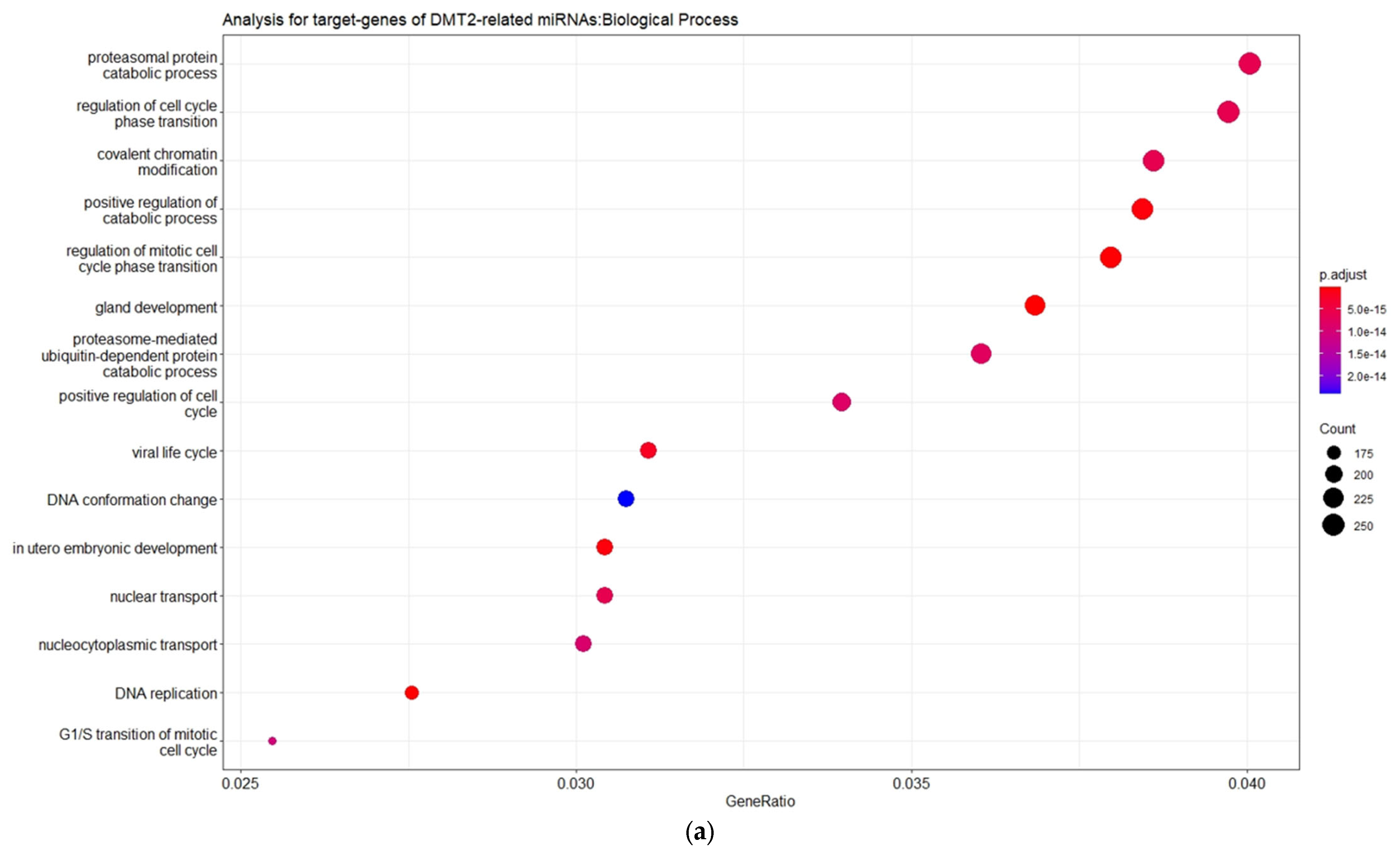
Table S2). The gene ontology enrichment analysis based on the biological process (BP) revealed that the miRNA-target genes are involved mainly in catabolic processes mediated by proteasome, and in processes related to cell cycle phase transition and nuclear transport (
Figure 3).
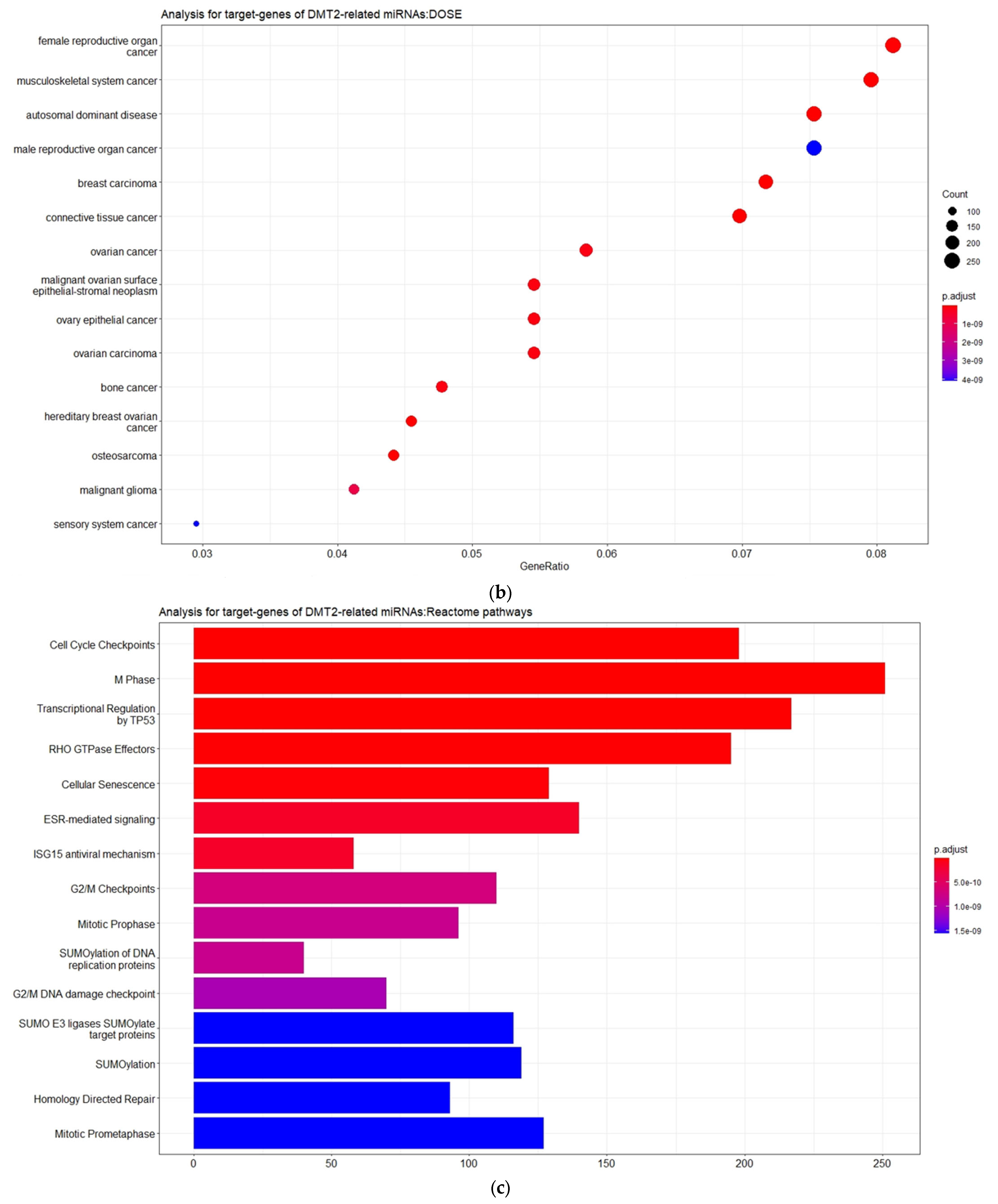
Moreover, as also shown in
Figure 3, the disease ontology (DOSE) enrichment analysis unveiled that the miRNA-target genes are implicated in different types of cancers, including tumors of the female reproductive system and the musculoskeletal system, breast cancer, and ovarian cancer. Although the association between DMT2 and cancer has been extensively reported and discussed over the last decades, the mechanisms underlying this relationship remain unclear [
28]. However, some epidemiological studies have revealed that DMT2 shares risk factors and characteristics with gynecologic breast, ovarian, and endometrial cancer, suggesting that lifestyle- and health-associated problems (e.g., obesity) are key factors for both illnesses [
29,
30,
31]. The hormonal changes of women with diabetes have been strongly associated with increased breast and ovarian cancer risk [
32,
33]. Indeed, the increased bioavailability of estrogen triggered the proliferation of ER-positive and/or estrogen-dependent breast tumor cells, while the activity of insulin and IGFs (insulin-like growth factors) has been also implicated in the positive proliferation and invasiveness of breast and ovarian cancer cells [
34,
35,
36]. In addition, DMT2 can affect the musculoskeletal system, leading to connective tissue alterations via metabolic changes such as protein glycosylation, blood vessel destruction, and nerve destruction, as well as collagen accumulation in skin and periarticular structures. However, a direct association of DMT2 and cancer in the musculoskeletal system has not been reported yet [
37].
Furthermore, the validated target genes (VTGs) of DMT2-related miRNAs are significantly enriched in pathways related with cell cycle regulation, transcription regulation by TP53 protein, and the SUMOylation processes that affect gene expression. Indeed, it was previously shown that deregulation of the post-translational modification process executed by the SUMOylation pathway affects insulin secretion in β-pancreatic cells [
38]. In addition, accumulated evidence during the last years proposes an important role of p53-signaling pathways in metabolic abnormalities (e.g., oxidative phosphorylation, glycolysis, lipolysis, lipogenesis, β-oxidation, gluconeogenesis, and glycogen synthesis) that result in the imbalance of insulin action [
39]. Moreover, insulin signaling has shown to regulate the FoxM1/PLK1/CENP-A pathway and, thus, promote the proliferation of β-pancreatic cells [
40].
3.3. Targeted Analysis to Reveal Unique Shared Genes between the Experimentally Validated Target Genes of DMT2-Related miRNAs and the Genes Implicated in DMT2 Pathogenesis Relevant to Pharmacogenomics Analysis
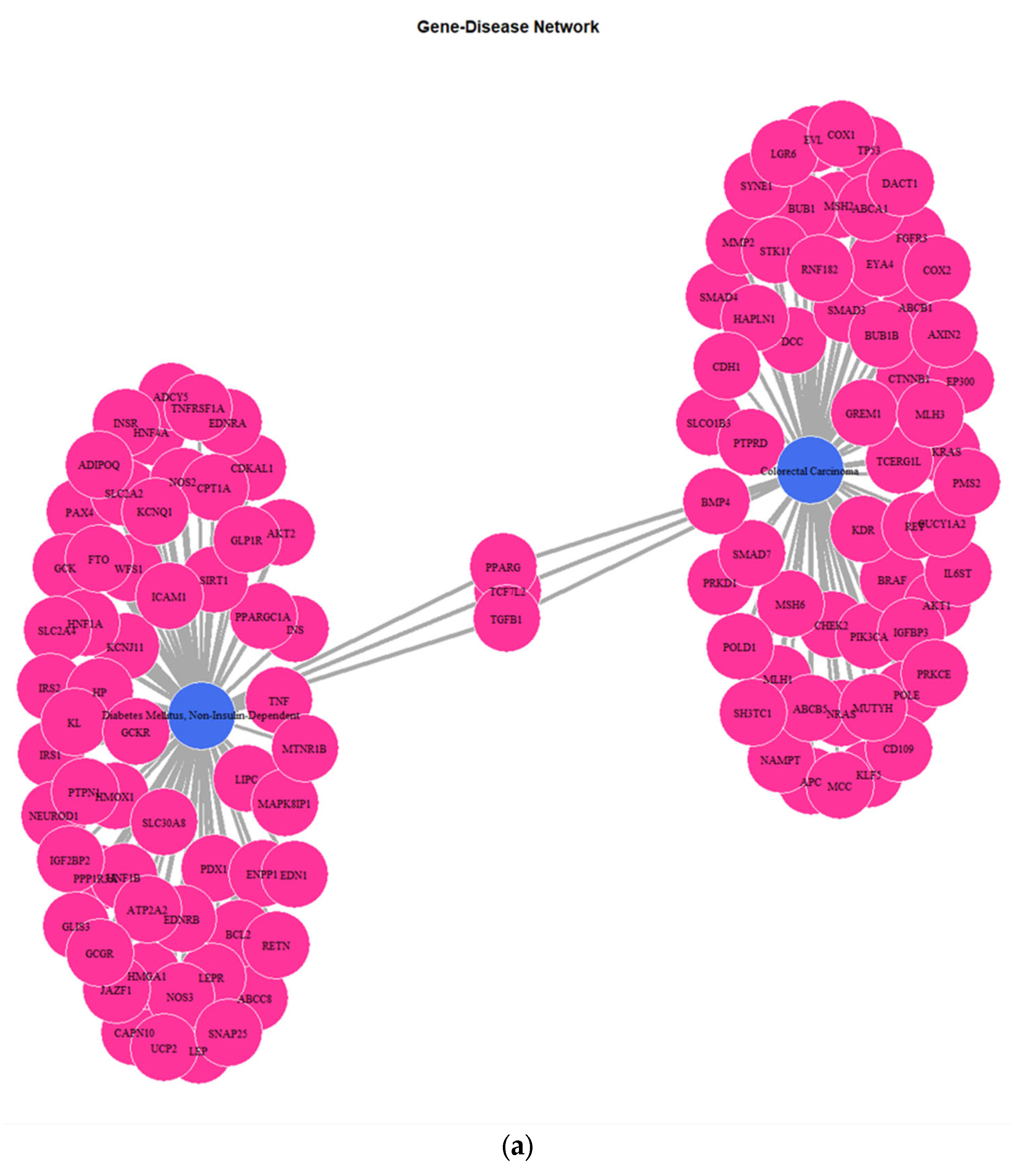
The correlation of 221 genes that are related to DMT2 pathogenesis, according to the DisGeNET database and the experimentally validated target genes (VDTs) from our analysis of the DMT2-associated miRNAs, revealed 71 unique shared genes (see
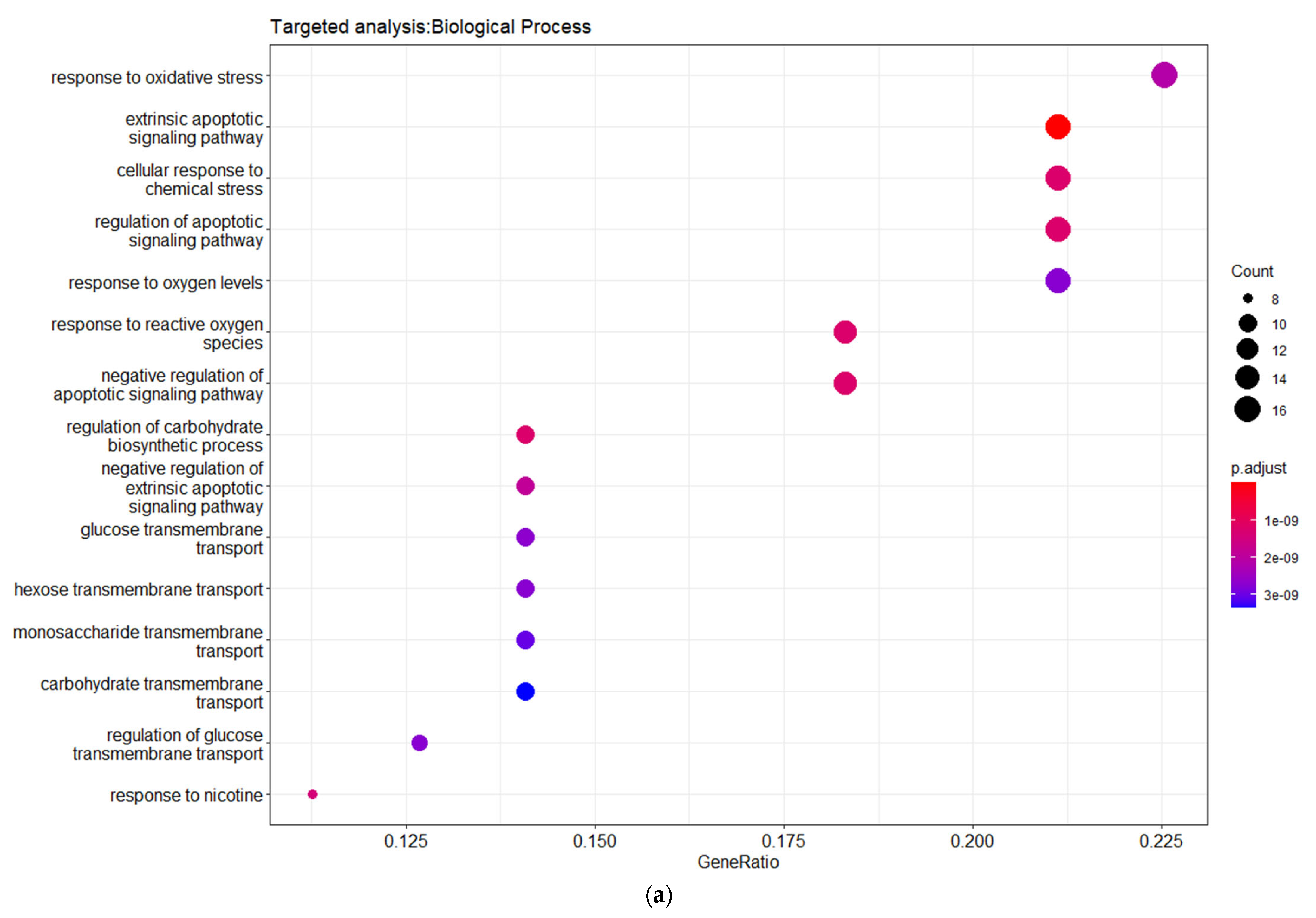
Table S3). These genes are related to DMT2 pathogenesis and interact with DMT2-associated miRNAs. The gene ontology enrichment analysis based on biological process (BP) revealed the shared genes involvement in processes related with oxidative stress, apoptosis regulation, and glucose transmembrane transport (
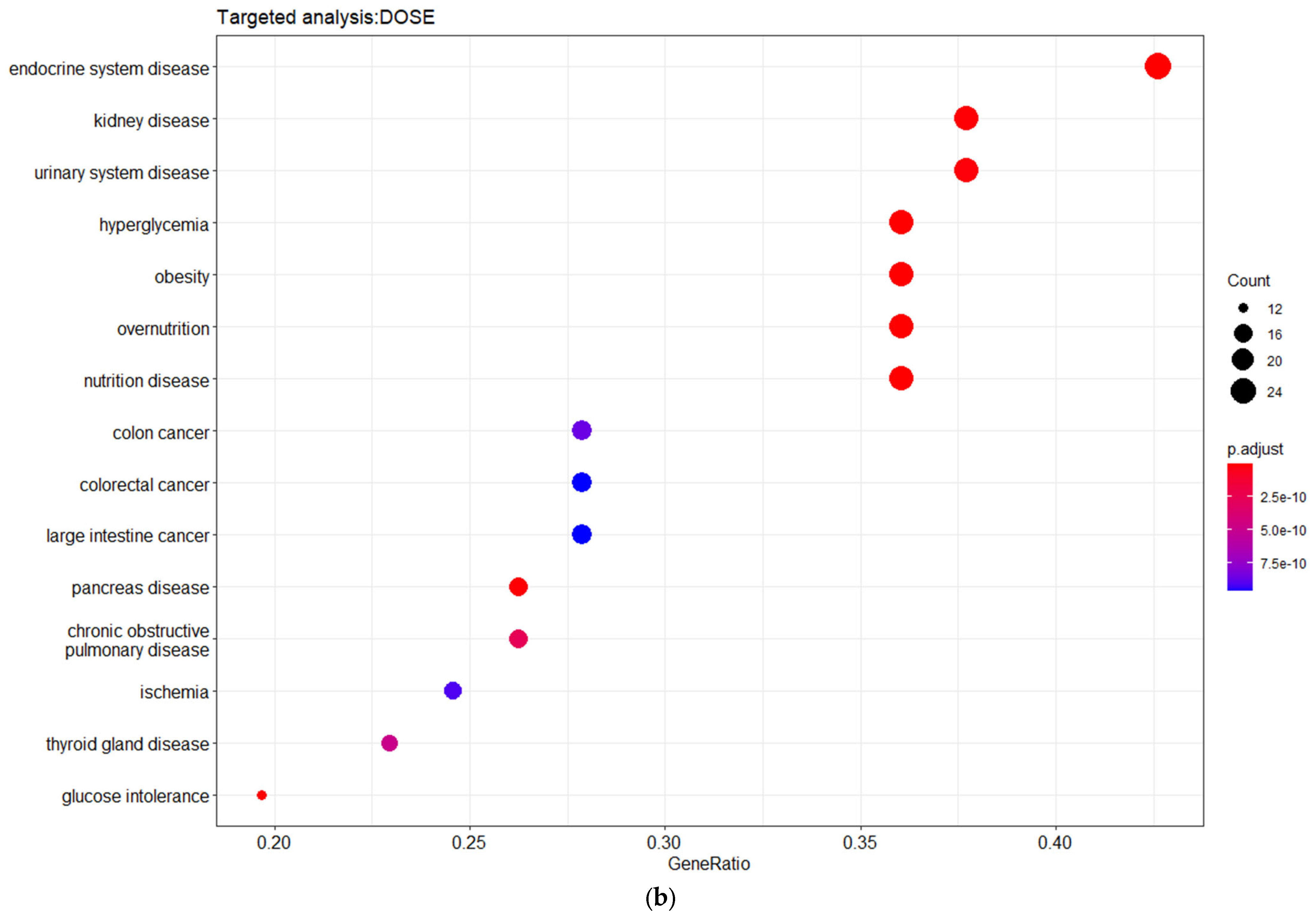
Figure 4). The disease ontology (DOSE) enrichment analysis unveiled that the shared genes are implicated in several diseases, which can be clustered in three major disease groups, including urinary system disease, obesity-overnutrition, and colorectal cancer (
Figure 4). The network (
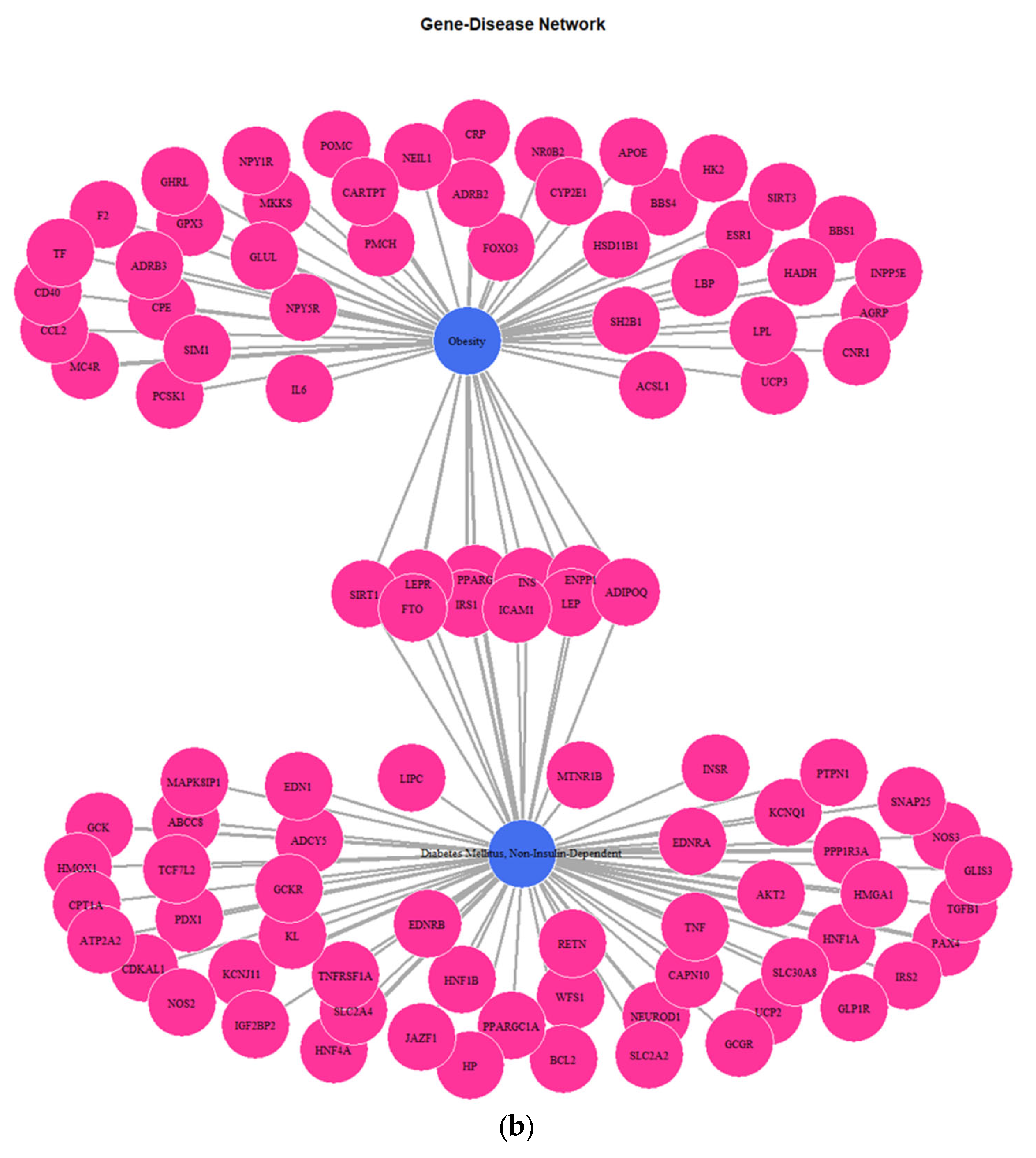
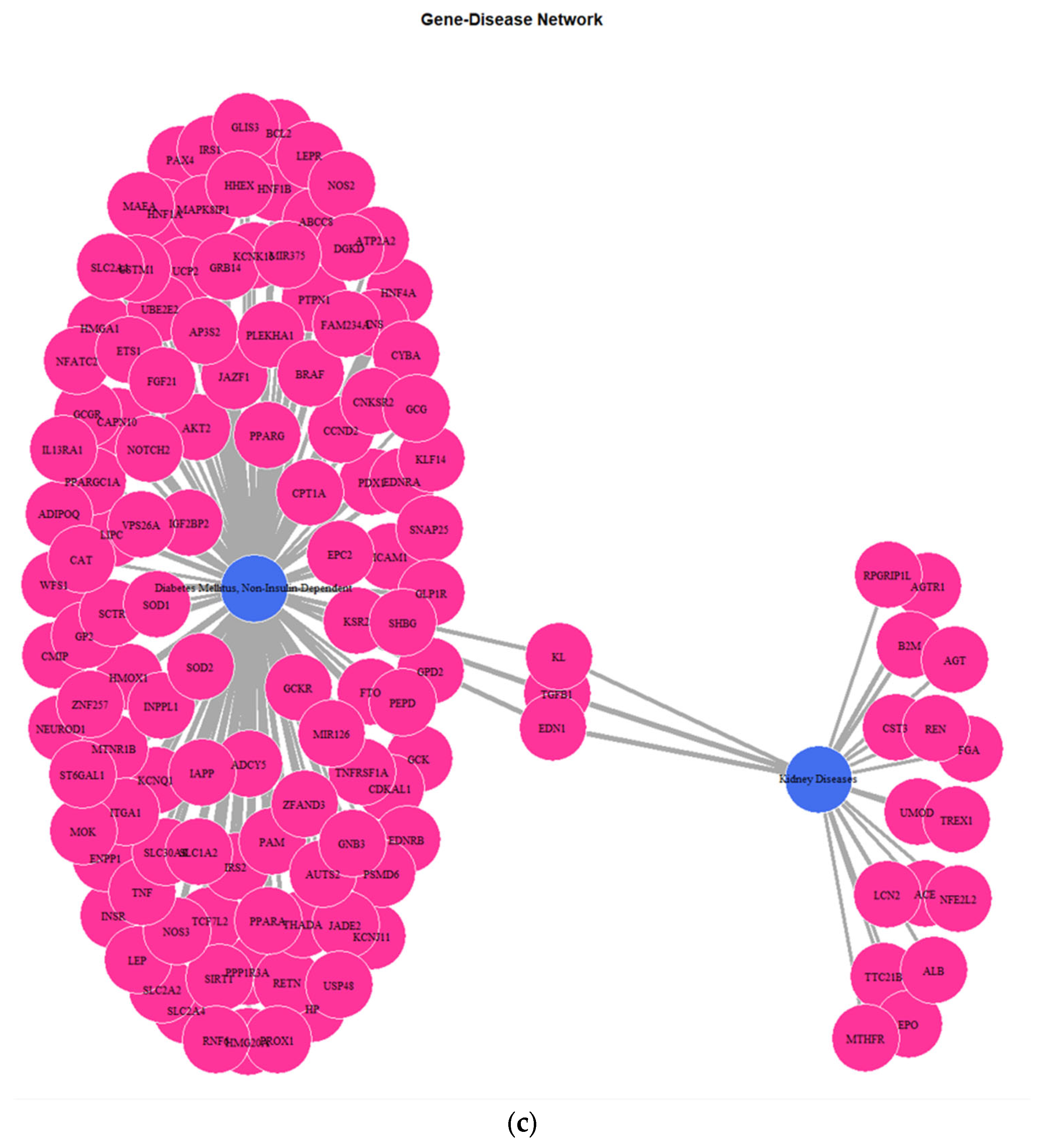
Figure 5) depicting the genes implicated in DMT2, kidney diseases, obesity, and colorectal cancer reveals several key genes shared among the disease groups. The shared key genes could be the connection link, in terms of pathophysiological mechanisms, which relates DMT2 with other diseases that coexist in patients with DMT2, including kidney diseases, obesity, and colorectal cancer. TGF-β1, a pleiotropic cytokine involved in several processes, including angiogenesis, extracellular matrix (ECM) formation, and immunoregulation, is considered a dominant factor involved in fibrotic processes of chronic kidney disease in patients with diabetes [
41]. The EDN1 gene encodes a preprotein that is subjected to the proteolytic process generating an endothelial cell-derived peptide (endothelin 1). Hyperglycemia and hypertension associated with DMT2 contribute to increased renal Endothelin 1 production, progressively resulting in diabetic nephropathy [
42,
43]. Several genes are shared between DMT2 and obesity-overnutrition, including the ICAM1, PPARG, LEPR, SIRT1, FTO etc. For instance, genetic variations in the FTO gene are related with insulin resistance, inflammation, obesity, and DMT2. Groups of patients with a high risk of genetic variations in the FTO gene could be benefited by obesity prevention programs to prevent DMT2 development [
44].
Furthermore, strong epidemiological evidence supports the existence of molecular connectors that increase the susceptibility for colorectal cancer (CRC) development of patients with DMT2. Diabetes is associated with dysregulated carbohydrate and lipid metabolic processes. The increased availability of nutrients, such as glucose and lipids, related with DMT2 disease favors the metabolic transformation of the cancer cells, overcoming growth inhibition checkpoints and apoptotic mechanisms [
45]. Several signal transduction pathways, including the Wnt, PI3K/Akt, Ras-ΜAPK, and TGF-β, are triggered by the high lipid and glucose content in patients with DMT2 [
46,
47]. Notable previous studies have demonstrated the increased activation of cancer-related pathways in non-cancer mucosa of DMT2 patients, suggesting the field cancerization effect induced by diabetes [
48]. TCF7L2, a gene shared between DMT2 and colorectal cancer, according to our analysis, is a transcription factor involved in the Wnt-signaling pathway [
49]. This gene is related with DMT2 development and its complications, as well as with a susceptibility to CRC evolution [
50]. Furthermore, previous studies have demonstrated a strong association between variants in the TCF7L2 gene and DMT2 and colorectal cancer development [
51,
52].
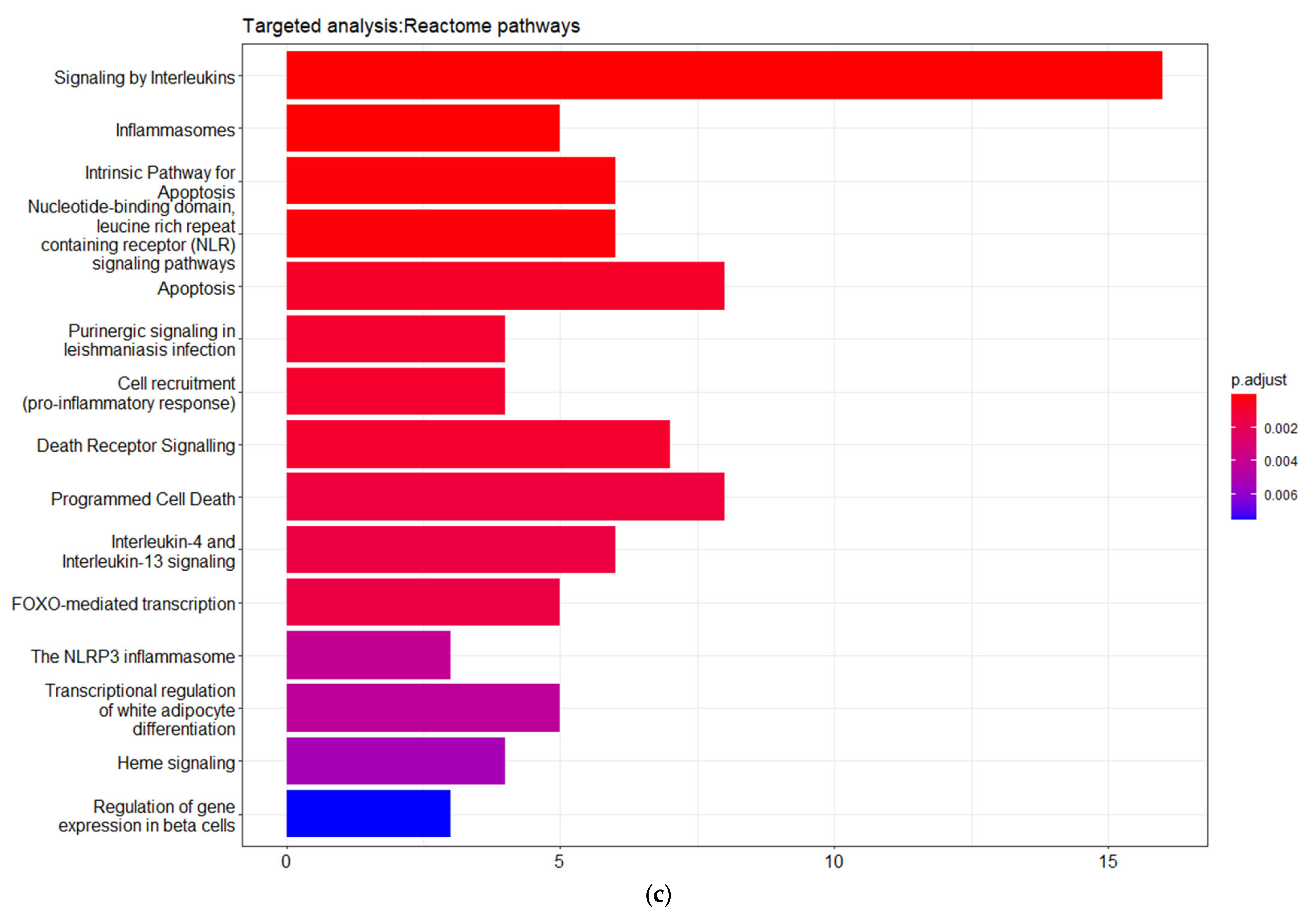
In addition, the matched genes are significantly enriched in pathways related with interleukins signaling, inflammasome and pro-inflammatory responses, apoptosis regulation, programmed cell death, and death receptor signaling. These pathways strongly interact with each other, forming larger pathway clusters as depicted on
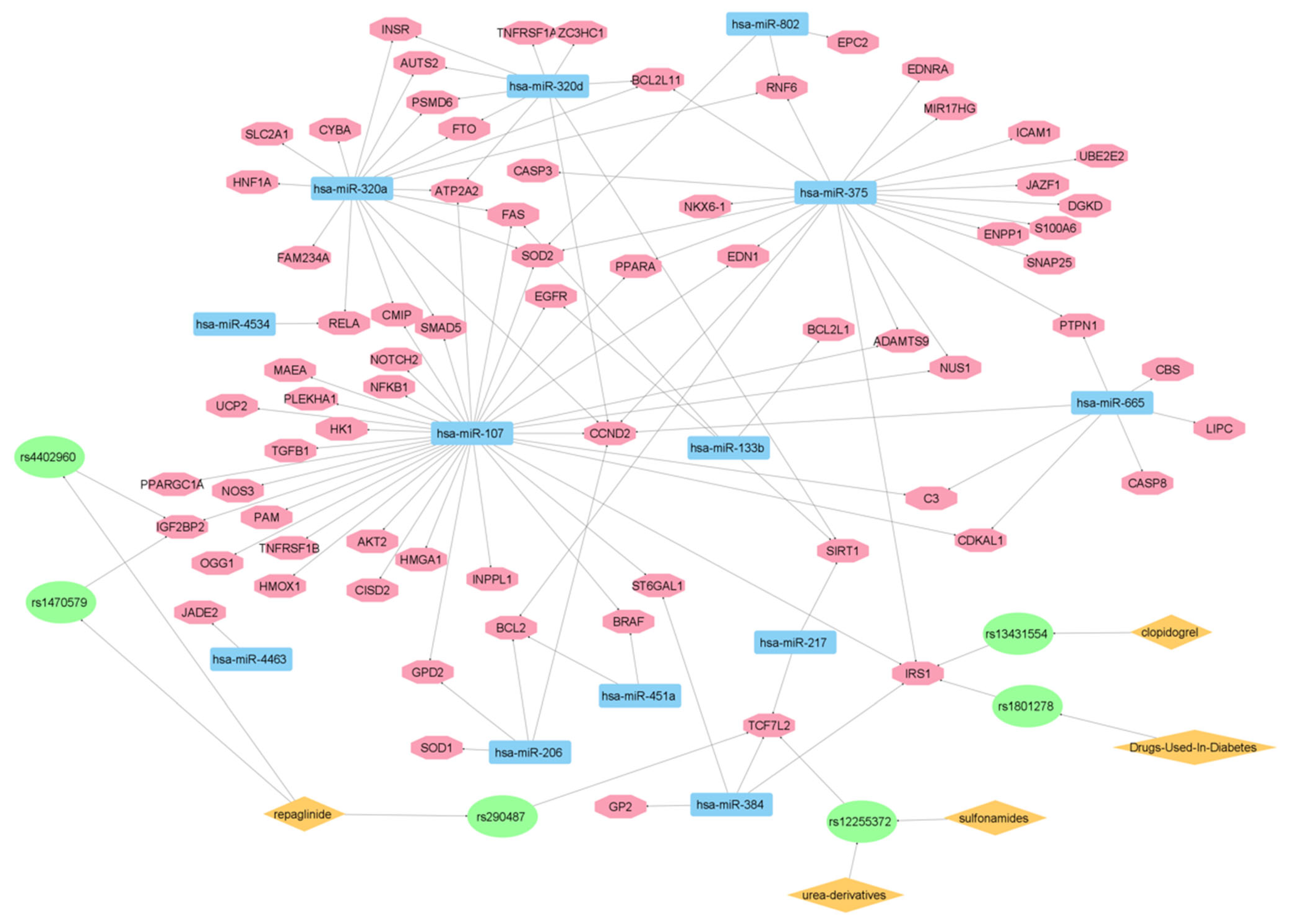
Figure 4d. Following these data, as shown in
Figure 6, an additional network analysis was carried out to link the identified genes and miRNAs implicated in DMT2 pathogenesis with drug targets. Importantly, in this network, 13 microRNAs (miR-802, miR-320a, miR-320d, miR-375, miR-665, miR-107, miR-133b, miR-4534, miR-4463, miR-451a, miR-217, miR-206, and miR-384) are connected via six gene single nucleotide polymorphism (SNP) variants (rs290487, rs4402960, rs1470579, rs12255372, rs1801278, and rs13431554) with DMT2 drug therapeutics. These SNPs are variants of insulin-like growth factor 2 mRNA-binding protein 2 (IGF2BP2) gene (rs4402960 and rs1470579) of transcription factor 7-like 2 (TCF7L2) gene (rs290487 and rs12255372) and of insulin receptor substrate-1 (IRS-1) gene (rs1801278 and rs13431554) [
53,
54,
55,
56].
Such a network allows the creation of connection molecular maps between genes, SNPs, microRNAs, and drugs (e.g., repaglinide, urea-derivatives) relevant to pharmacogenomics and precision medicine decisions in DMT2 therapy. It is also interesting to note that one SNP (rs13431554) is associated with clopidogrel effects sharing the common target gene insulin receptor substrate 1 (IRS1) with the DMT2-associated SNP rs1801278. The latter correlation was shown to exist in coronary artery disease (CAD) patients with DMT2, where IRS1 polymorphisms and high platelet reactivity were associated with the clopidogrel therapy outcome [
56].
Notably, IRS1 is also known as a signaling adapter protein, which is to be involved in various signaling cascades with a potential role in cancer progression [
57,
58]. Such connections allow us to better understand how specific gene functions contribute to the molecular pathophysiology of various illnesses. Recently, researchers proved that miR-107 expression is regulated by the p53 pathway, leading to insulin metabolism dysfunction [
59]. Moreover, miR-133a plays a key role for proper skeletal and cardiac muscle function by regulating crucial signaling pathways, including the Wnt signaling [
60]. It was also shown to regulate the EGFR/cMyc/P53 axis. Thus, it is implicated in various cellular processes, including migration, invasion, autophagy, proliferation, and apoptosis [
61]. Since the function of miRNA-133a was previously associated with diabetic cardiomyopathy [
62], it is reasonable for someone to consider molecular connections from its pleotropic biological effects with DMT2 pathophysiology.
Likewise, for microRNA-375, it was previously published that it exerts important gene functions that affect the complex regulatory network of pancreatic development [
63]. Besides cell growth and proliferation, it has been proposed that its function in β-pancreatic cells is correlated with insulin secretion [
64]. Complementary previous studies have also shown that miR-375 is involved in the regulation of the epithelial–mesenchymal transition (EMT) of tumor cells by affecting signaling pathways like Wnt, nuclear factor κB (NF-κB), and transforming growth factor β (TGF-β), which are considered crucial for cancer cell progression [
65].
As far as the function of miR-320 in DMT2 is concerned, its involvement in the regulation of glucose and lipid metabolism was previously proposed. Specifically, the diabetes-induced cardiac dysfunction was shown to be mediated by miR-320 via the transcription regulation of genes involved in the fatty acid biosynthetic pathway [
66]. Furthermore, miR-320a can directly cause dysfunction of pancreatic β-cells via its target gene [
67], whereas it has been proposed to exert effects to myelodysplastic syndromes, too [
68].
Overall, the proposed network analysis that unifies knowledge from genes, miRNAs, SNPs, and drugs sheds light onto molecular maps that contribute to: (a) Better understanding of DMT2 pathophysiology; (b) The creation of connections with signaling pathways linked to other illnesses; (c) The work related to drug repurposing approaches; (d) The exploitation and clinical implementation of pharmacogenomics knowledge; and (e) The formulation of precision medicine decisions in DMT2 therapy.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

















