1. Introduction
It is well-known that the develo** nervous system presents higher sensitivity to some chemical exposure when compared to the adult nervous system. This fact occurs because normal development of the nervous system commonly requires spatial and temporal coordination of critical events, such as proliferation, migration, differentiation, synaptogenesis, myelination, and apoptosis, that can be impaired by toxicants [
1,
2]. Despite this, developmental neurotoxicity (DNT) evaluation is not mandatory in the registration of substances for regulatory purposes, which means these evaluations are only conducted when relevant evidence is pointed out by previous reports [
3]. Currently, guidelines to determine the potential for DNT are basically provided by the United States Environmental Protection Agency (U.S. EPA) Office of Prevention, Pesticides, and Toxic Substances (OPPTS) 8706.300 and the Organization for Economic Co-operation and Development (OECD) Test Guideline no. 426 [
4,
5]. In general, both guidelines involve a long test series, performed preferably in rats, that includes behavioral, neuropathological, and motor activity evaluation, as well as sensory function and learning and memory testing. However, the large number of animals, long duration of tests, and intensive use of resources have been raising criticism and drawing attention to the need for more realistic test protocols for regulatory purposes, especially in terms of cost-effectiveness [
6,
7,
8].
Various in vitro cell culture methods for assessing DNT have been used, and they include mainly rodents and human cell lines [
9,
10,
11,
12]. Despite the clear advantages of using cell culture techniques, e.g., consistency, reproducibility, and control of the physicochemical environment, there is no doubt that one of the biggest challenges is faithfully reproducing the complexity of a develo** nervous system. Hence, alternative methods based on whole organisms have been said to be more suitable to fill these gaps, in addition to the fact that they used to provide refinement or reduction of animal use [
13]. Consistent with this idea, Selderslaghs et al. [
14] suggested a new alternative method for DNT testing based on tail coilings in zebrafish (
Danio rerio) embryos, thus demonstrating later that a considerable majority of known, tested developmental neurotoxicants resulted in hypo- or hyperactivity in exposed embryos [
15]. DNT investigation using tail coiling assay was further assessed after exposure to a variety of chemicals, such as pesticides [
16,
17,
18,
19,
20], ethanol [
21], flame retardants [
22,
23], cannabidiol [
24], and other compounds [
25,
26], evidencing the potential of the method as well as the model system.
The zebrafish is a vertebrate organism that offers several advantages. Whereas development stages are difficult to assess in most mammalian models in toxicology studies, the external fertilization and transparency of zebrafish embryos allow monitoring of a well-known development from the first cell cleavage [
27]. Easy handling and selective permeability of chorion facilitate egg exposure, while the rapid development—the embryonic stage lasts about 72 h post-fertilization (hpf)—makes the assessment of multiple endpoints in a short time feasible [
28,
29]. This characteristic is especially crucial to the suitable conducting and interpretation of data generated by embryos’ tail coiling assays. Spontaneous tail coiling in zebrafish involves side-to-side contractions of the trunk and consists of the first motor behavior of embryos. It begins at 17 hpf, reaching a peak at 19 hpf, and then decreases gradually, resulting in a different motor activity known as “touch-induced response” by 26 hpf [
30]. Thus, it should be expected that the analysis of tail coiling activity must be performed in a period of development where no significant variations in coiling activity are observed, in order to avoid misinterpretation of results.
Over the last decade, valuable contributions have been added to the tail coiling assay. The potential of coiling activity to discriminate modes of action for compounds interfering with neurotransmission was firstly investigated by Vliet et al. [
31]. Later, Zindler et al. [
32] correlated effects triggered by known neurotoxicity compounds on coiling activity with sublethal effects in develo** embryos. Time-saving was also enhanced by combining the tail coiling assay with high-content screening (HCS) [
33]. Zindler et al. [
34] evidenced the influence of dark/light transition during tail coiling assay, also demonstrating that non-teratogenic concentrations of ethanol can delay the onset of coiling activity in exposed embryos. More recently, Ogungbemi et al. [
35] optimized important experimental parameters, such as embryonic developmental stage (23 to 24 hpf), sample size (20 embryos per video), rearing conditions, exposure, acclimation, and analysis duration. They also found that there is no influence between adjacent embryo activity, allowing them to analyze a group of embryos simultaneously. Despite the efforts for standardization of the method, there is no consensus related to the most suitable age of embryos, and therefore, different developmental stages in which coiling activity is to be assessed have been proposed. Nevertheless, it is crucial to highlight the need for prioritizing rapid and efficient protocols for the detection of toxic effects as early as possible.
In DNT studies, chemicals with known effects are often used as positive controls to demonstrate the validity of a method, and verify the sensibility of parameters to be used as well. To be a suitable positive control, the chemical must trigger an effect that can be measured by the method under development. Therefore, a suitable positive control to be used in the tail coiling assay for DNT evaluation needs to affect the coiling activity, either by increasing or decreasing the motor activity of exposed embryos [
36,
37].
Toxicity testing of substances commonly requires organic solvents for complete solubilization and toxicant delivery at effective concentrations. In zebrafish, the most frequently applied solvent is dimethyl sulfoxide (DMSO). Other solvents can be required when sufficient solubility is not obtained using DMSO [
38]. For this reason, it is crucial to establish safe concentrations of these compounds for a tail coiling assay, since the central nervous system is known to be a target of many organic solvents [
39].
In order to contribute experimentally to a harmonized protocol for tail coiling assay and facilitate the comparability of results, the present study aimed to (i) monitor early coiling activity and throughout embryonic development, to select the most appropriate stage of development for a fast protocol; (ii) test substances with known mechanisms of action as candidates for positive controls; and (iii) establish safer concentrations of widely used organic solvents for tail coiling assay.
2. Materials and Methods
2.1. Chemicals and Test Solutions
Caffeine (purity = 98.5%) was obtained from Acros Organics (Geel, Belgium). Fluoxetine hydrochloride (purity > 98%) was purchased from Tokyo Chemical Industry (Tokyo, Japan). Tricaine methanesulfonate (MS-222; purity = 98%), ethanol (purity ≥ 99.5%) and DMSO (purity ≥ 99.5%) were purchased from Sigma-Aldrich (St. Louis, MO, United States). Acetone (purity = 99.6%) was purchased from J.T. Baker (Phillipsburg, NJ, United States). Stock solutions of caffeine (15 mg/mL) and MS-222 (10 mg/mL) were prepared by dissolving the powder in the embryo medium (2 mM CaCl
2, 0.5 mM MgSO
4, 0.75 mM NaHCO
3, and 0.07 mM KCl in ultrapure water according to ISO 7346-1) [
40]. For obtaining test solutions, stock solutions were diluted in embryo medium just before the experiments. Solutions containing MS-222 had no significant changes in their pH at tested concentrations. A stock solution of fluoxetine (1 mg/mL) was prepared in DMSO and diluted to the final concentrations in embryo medium. In all fluoxetine solutions, DMSO concentration did not exceed 0.01% (
v/
v), as recommended by the Organization for Economic Cooperation and Development (OECD) test guideline no. 236 [
41]. Acetone, DMSO, and ethanol were prepared directly in test solutions, which were also obtained fresh each day.
Table 1 provides the concentrations tested for each substance.
2.2. Zebrafish Husbandry and Egg Acquisition
All embryos used in the following experiments were provided by the Zebrafish Facility at the Department of Clinical Analyses, Toxicology and Food Sciences from the School of Pharmaceutical Sciences of Ribeirão-Preto/University of São Paulo. Male and female adult zebrafish were maintained in a flow-through system (ZebTEC, Tecniplast, Italy), with a maximum density of 20 fish per tank. The water of the system was set at 26 ± 1 °C, with pH ranging from 7.25 to 7.35 and conductivity between 290 and 320 μS. Lighting was provided by a constant cycle of 13 h light/11 h dark (lights on at 8:30 a.m.), and fish were fed twice a day with commercial flake feed (Tetra) and
Artemia sp. eggs (Sera). The day before egg acquisition, males and females (2:1) were transferred and kept separated overnight into 1 L breeding tanks. Once the light started in the morning, the divider was removed, and eggs were collected about 90 min later. By using a stereomicroscope, viable eggs were established according to Kimmel et al. [
29] and randomly selected for subsequent experiments.
2.3. Test Design
A total of three potential candidates for positive controls of the tail coiling assay were selected: caffeine, fluoxetine, and MS-222. For testing the influence of organic solvents, acetone, DMSO, and ethanol were chosen. In all experiments, embryos up to the late blastula period (sphere stage, 4 hpf) were transferred to six-well plates (20 embryos per well) filled with 10 mL of embryo medium, fresh test solutions, or their respective control solutions (embryo medium for MS-222; caffeine, acetone, ethanol, and DMSO solutions; DMSO 0.01% v/v for fluoxetine solutions). Immediately, plates were incubated at 27.5 ± 0.5 °C with a photoperiod of 13h light/11h dark. For protection from evaporation, plates containing acetone and ethanol solutions were sealed with Parafilm-M. For MS-222 exposure, embryos were initially kept in water, and at 24 hpf (2 h before the experiment), the water was replaced by the same volume of MS-222 test solution. The respective embryos in the control groups underwent the same procedure. Experiments to determine the stage of development, the efficiency of positive controls, and effects of organic solvents on the tail coiling assay were conducted separately using embryos from different batches. Also, independent experiments containing embryos from at least two couples each were used per treatment.
2.4. Tail Coiling Assay
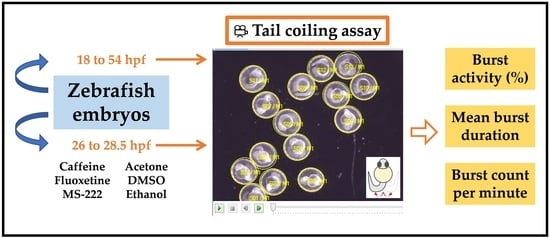
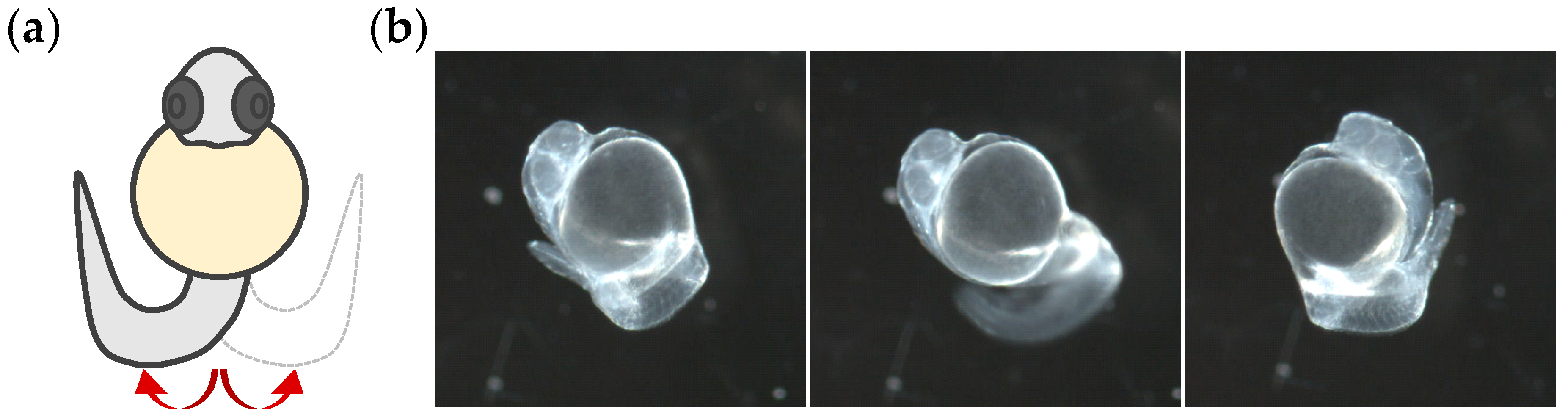
Non-dechorionated embryos checked for morphological abnormalities were transferred to single concave glass slides (75 mm × 25 mm; 20 embryos per cavity). To monitor the coiling activity throughout development, embryos from 18 to 54 hpf were videotaped every 2 h. Then, to shorten the time between measurements and investigate critical periods of embryonic development, new monitoring of coiling activity from 22 to 30 hpf at hourly intervals was conducted. To minimize the inconvenience of removing the embryos from the incubator every 1 or 2 h, different groups of 20 embryos were used per time. For positive control candidates and treatments with organic solvents, coiling activity was monitored from 26.0 to 28.5 hpf. The temperature and lighting of the room were kept constant, and 5 min were defined for acclimation to minimize disturbance during the transfer of embryos. Tail coiling activity was recorded for 3 min using a camera (AxioCam ICc 5, Zeiss, Germany) coupled to a stereomicroscope (Stemi 508, Carl Zeiss, Germany) with darkfield illumination. During the video monitoring, embryos did not receive any sensory stimulation. Movies were obtained at a frame rate of 10 frames per second (fps) and processed in Audio/Video Interleaved (AVI) (Motion-JPEG compression) format. Raw data were extracted and analyzed by DanioScope software (Noldus, version: 1.1, Wageningen, The Netherlands). To define the region of interest, a contour was drawn over the chorion, and when floating occurred, the adjustment was limited to a maximum of five measurements per embryo. Coiling activity was reported based on the following parameters: percentage of time the embryo was moving (“burst activity”), percentage of time of inactivity (“inactivity”), the sum of all movement durations (“mean burst duration”), and the number of embryo movements per minute (“burst count per minute”).
Figure 1 shows a representative image of the tail coiling activity exhibited by an embryo at 26 hpf.
2.5. Statistical Analysis
Normality was tested with the Kolmogorov–Smirnov test. As the majority of data did not present a normal distribution, differences between groups were checked by ANOVA on ranks (Kruskal–Wallis test), followed by the Dunn’s post-hoc test. Data obtained from monitoring coiling activity throughout embryonic development were tested to significant differences against each other. Treatments with caffeine, fluoxetine, MS-222, and organic solvents were compared with their respective controls. All values are shown as mean and 95% confidence intervals. Statistical analysis was carried out using GraphPad Prism (version: 5.1). Results were defined as significant for a value of p ≤ 0.05.
4. Discussion
This study provides contributions to screening for DNTs through the establishment of experimental parameters for an alternative testing method, based on the tail coiling activity of zebrafish embryos. For the first time, tail coiling activity was monitored throughout development only 1 h after the onset of motor activity (as mentioned before, the first motor behavior of zebrafish embryos occurs at 17 hpf), until the hatching period at 54 hpf. Embryos were monitored every 2 h for the frequency (“burst count”) and duration (“mean burst duration”) of coils. We also reported tail coiling activity through the percentage of time spent in activity (“burst activity”), and the percentage of time the embryo was immobile (“inactivity”), which was particularly useful for monitoring the hypoactivity. Furthermore, we tested candidates for positive controls able to induce alterations in all parameters of coiling activity. Finally, we have found safe concentrations of widely used solvents in zebrafish experiments, and demonstrated which one of them should be avoided in the tail coiling assay.
Motor behavior in zebrafish embryos is divided into two types of patterns: spontaneous tail coiling and touch-induced response [
30]. Spontaneous tail coiling results from an active neural network in the still-immature spinal cord, and is based on electrical coupling [
42]. The second pattern of motor behavior appeared at 21 hpf, when the contractions were mediated by neurotransmitters, resulting in faster and stronger contractions than the first type of movement. As its name suggests, embryos also become sensitive to sensory stimulation [
43,
44]. Later studies have been able to distinguish a single coil—described as a single contraction of the trunk—from a double coil, characterized by two contralateral contractions of the trunk. Started at 24 hpf, double coils seem to represent most of the events by 27 hpf [
45]. Despite these alternating patterns of motor activity, our data showed a peak by 20 hpf, followed by a period of relative stability in the coiling activity of embryos. This pattern of motor activity is in agreement with the literature for dechorionated embryos, although we did not monitor coiling activity at 19 hpf, when the maximum intensity of spontaneous movements has been described to occur [
30,
46]. In a recent report, Ogungbemi et al. [
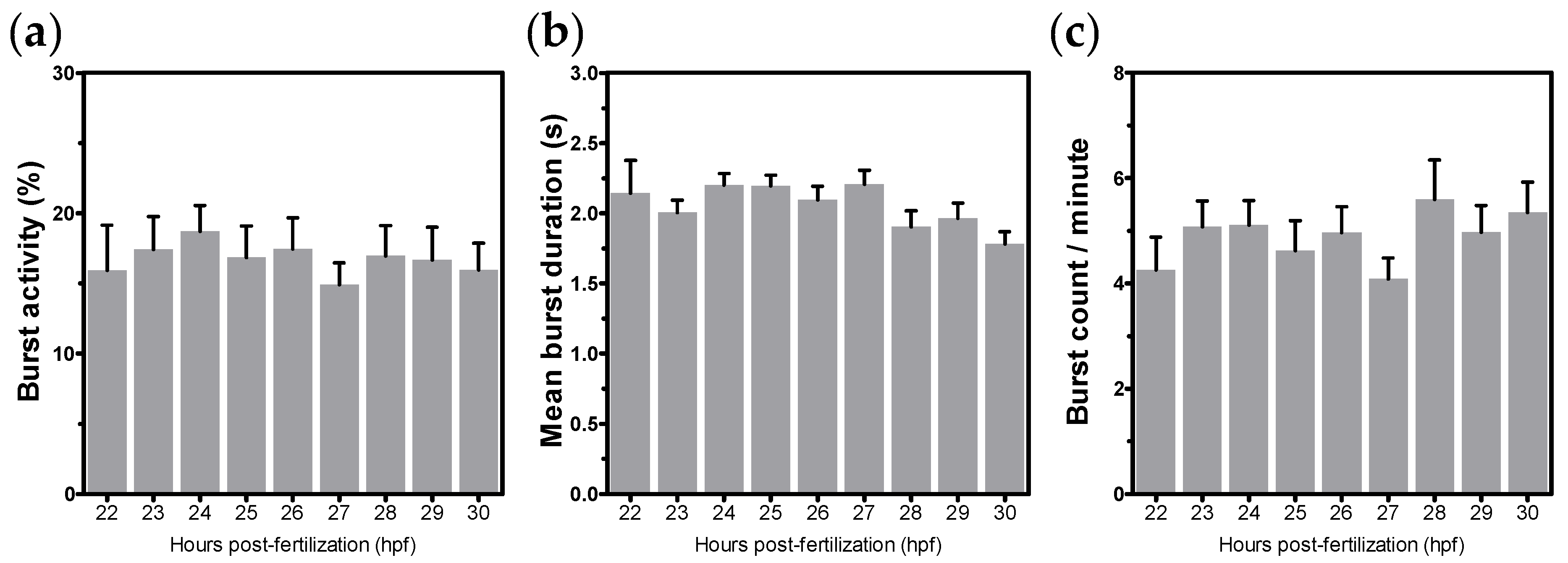
35] observed that there are no differences in the duration of acclimation period. They also demonstrated an absence of influence between adjacent embryos on tail coiling activity, allowing analyses of a group of embryos simultaneously. Sample size was also analyzed with up 20 embryos per video. These conditions are in accordance with the chosen parameters in the present work. Nevertheless, the peak of coiling activity was described by these authors as 23 and 24 hpf. Because of this intrinsic variability, the characterization of coiling activity has been recommended for each zebrafish strain to be used.
The embryonic development stage has been demonstrated to be crucial for tail coiling activity assessment. However, there is a considerable variability related to the age of embryos among studies. In Selderslaghs et al. [
14], the tail coiling activity was evaluated from 24 to 26 hpf. Most studies also conducted their measurements in embryos aged 24 hpf [
17,
18,
21,
24,
25], while others have used embryos at either 26 hpf [
19] or in the subsequent stages [
32,
34]. Less often, tail coiling activity has been monitored as early as 17, 18, or 19 hpf, and recorded at 1 or 2 h intervals until 25, 27, or 28 hpf [
22,
26,
47,
48]. Monitoring of the coiling activity after exposure to chemicals for a wider period range of development offers the possibility to investigate the effects of substances on different patterns of motor behavior. Despite that, given the time-consuming analysis, these protocols do not meet the requirement to be a fast protocol for DNT investigation. In our study, we found a period of relative stability, with no significant differences between measurements from 22 to 30 hpf in all parameters of coiling activity. However, because most spontaneous tail coiling events are expected to cease around 26 hpf, we choose this period of development and then conducted our analysis upon chemical exposure within the next 2.5 h. Then, as we initially demonstrated that no significant differences were found among measurements of the coiling activity in untreated embryos from 26.0 to 28.5 hpf (see hourly intervals measurements from 26 hpf to 29 hpf in
Figure 3), we assumed that significant alterations within this development stage would be a result of external factors, like exposure to chemicals.
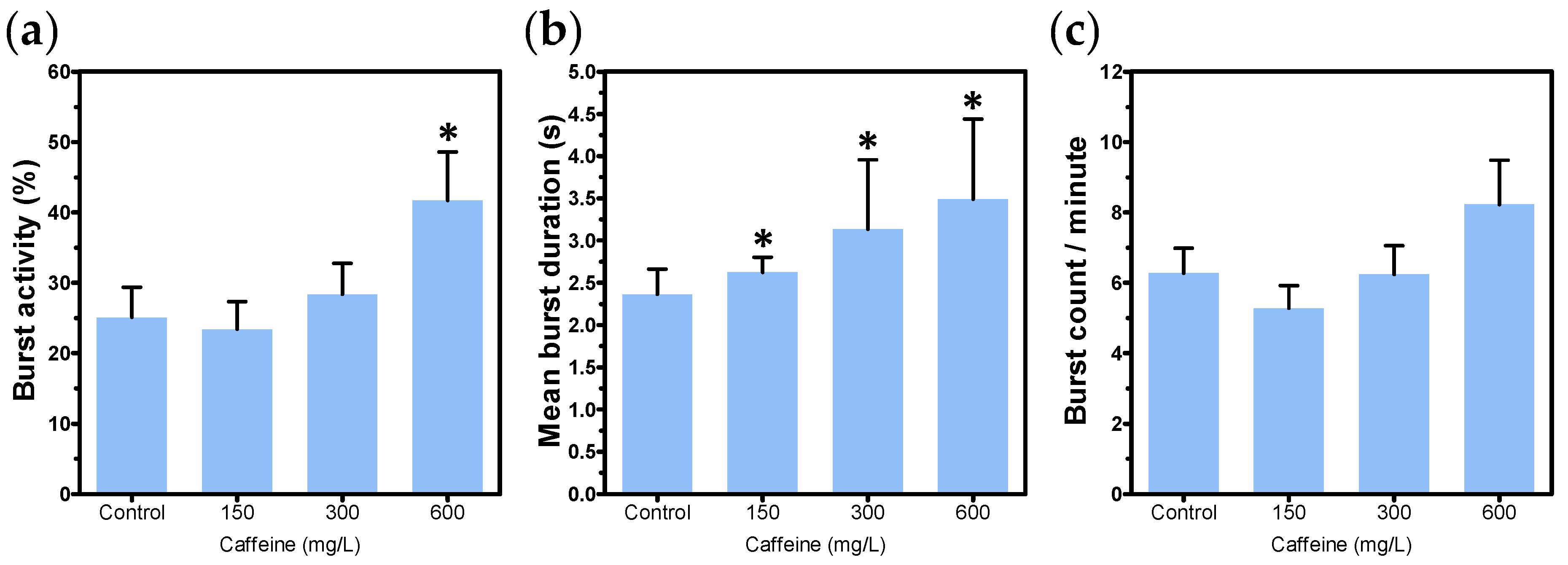
The selected positive control candidates for tail coiling assay induced either hyper- or hypoactivity. Caffeine is a stimulant of the central nervous system, mainly known to interfere in the mobilization of intracellular calcium, inhibit phosphodiesterases, and act as an antagonist in adenosine receptors [
49]. The motor stimulant action of caffeine has been reported in previous studies, and seems to be related to its effects on adenosine receptors [
50]. In zebrafish, exposure to caffeine has caused a higher frequency of spontaneous movements at lower concentrations (2.5 to 5.0 mM, or 485.5 to 970.9 mg/L), but stopped movements at the highest concentrations (10 mM or 1941.9 mg/L) [
51]. This ability of caffeine to produce opposite effects when tested in low or higher concentrations has already been described as a “biphasic profile”, and it has also been observed in rodents [
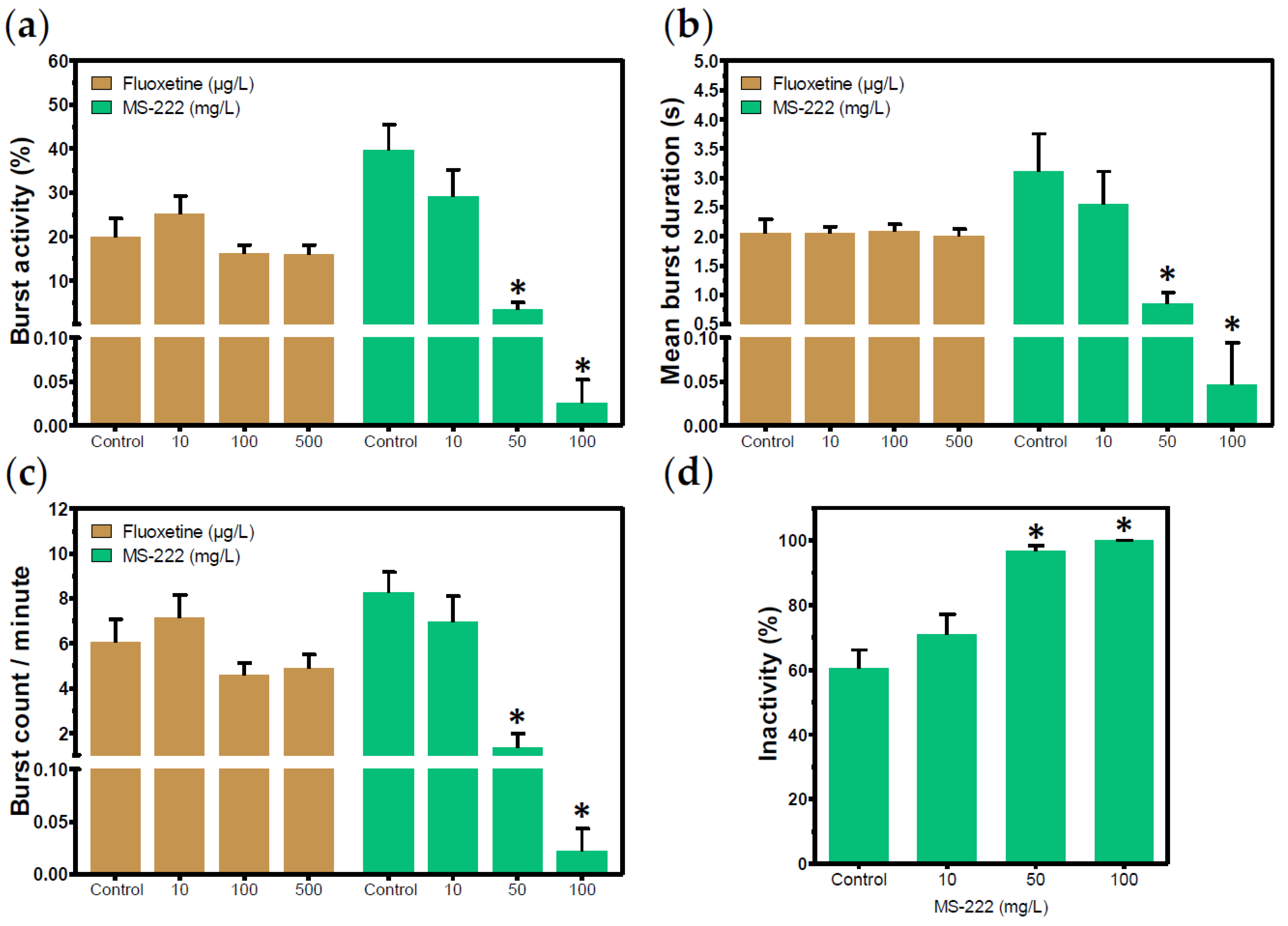
50]. In the present study, caffeine could prolong mean burst duration, thus resulting in a greater amount of time the embryo spends in movement when compared to the control group. These findings evidence the potential of caffeine as a positive control of hyperactivity in the tail coiling assay. However, in case the coiling activity is assessed at later stages of development, it is important to make sure that the selected concentrations are not able to induce morphological abnormalities that can impair the interpretation of results. Fluoxetine belongs to the group of selective serotonin reuptake inhibitors (SSRIs), and is commonly used in the treatment of psychiatric disorders [
52]. In studies with zebrafish, fluoxetine has been found to exert a decrease in locomotor activity of larvae [
53,
54], whereas no effects have been reported in either the spontaneous or touch-induced coiling of embryos [
55]. Following 22 h exposure to low concentrations of fluoxetine, our results show that the SSRI did not induce hypoactivity in exposed embryos, despite a slight tendency to decrease coiling activity at 100 and 500 µg/L (
Figure 5a,b). This fact was corroborated by Zindler et al. [
34], who only observed the absence of spontaneous movements after exposure to a teratogenic concentration of fluoxetine (48 mg/L), but not at concentrations in which embryos did not present sub-lethal effects until one day post-fertilization (dpf). On the other hand, MS-222 proved to be a promising candidate for positive control of hypoactivity in the tail coiling assay. MS-222 is a commonly used drug for anesthesia, sedation, and euthanasia of various fish species. Once absorbed through the gills or skin, MS-222 acts by blocking the entrance of sodium ions (Na+) into excitable cells, thus inhibiting action potentials [
56]. Here, we successfully abolished the coiling activity by using MS-222 100 mg/L, and a significant decrease was observed in embryos exposed to MS-222 50 mg/L compared to the control group. Similar responses in the coiling activity of zebrafish embryos were already reported after exposure to MS-222 [
57].
Toxicity induced by organic solvents to the early life stages of zebrafish has already been investigated [
38,
58,
59]. In this study, the exposure of embryos to organic solvents ranging from 0.01% to 1.00% allows us to find apparently safe concentrations for their use in the tail coiling assay. Concerning acetone, it is not yet clear why hyperactivity has been induced after exposure to the solvent. The anticonvulsant properties of acetone have been well-documented in the literature [
60]. Despite this, drugs that share the same effect have reduced spontaneous movements in zebrafish larvae, instead of inducing an increase in motor behavior [
61]. Due to its low toxicity and ability to solubilize different compounds, DMSO is by far the most extensively used solvent in zebrafish toxicity experiments [
38]. Our data suggest that even the highest concentration of DMSO (1%
v/
v) can be chosen for the tail coiling assay, since it did not cause alterations that could be detected in the activity of embryos. Even so, because DMSO can interfere in chorion permeability, it is suggested to work with the lowest concentrations, whenever possible, such as 0.01% [
62]. Among the solvents tested, only treatment with ethanol induced both hyper- and hypoactivity. Interestingly, a contradictory effect of ethanol was recently reported when the frequency of coils was monitored at different stages of development, inducing hypoactivity at 24 hpf but elevating coiling frequency in a concentration-dependent manner when analyzed at 26 hpf [
32]. Because of the effects induced by ethanol in the coiling activity of embryos at concentrations tested here, it may not be the solvent of choice. However, the potential of ethanol to induce hyper- and hypoactivity could be explored as a positive control in the tail coiling assay.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}