
Perspectives in Genome-Editing Techniques for Livestock


Abstract
:Simple Summary
Abstract
1. Introduction
2. Precision Genome-Editing Tools Successfully Used in Livestock
2.1. Zinc Finger Nucleases (ZFNs)
2.2. Transcription Activator-like Effector Nucleases (TALENs)
2.3. Clustered Regularly Interspaced Palindromic Repeats (CRISPR)
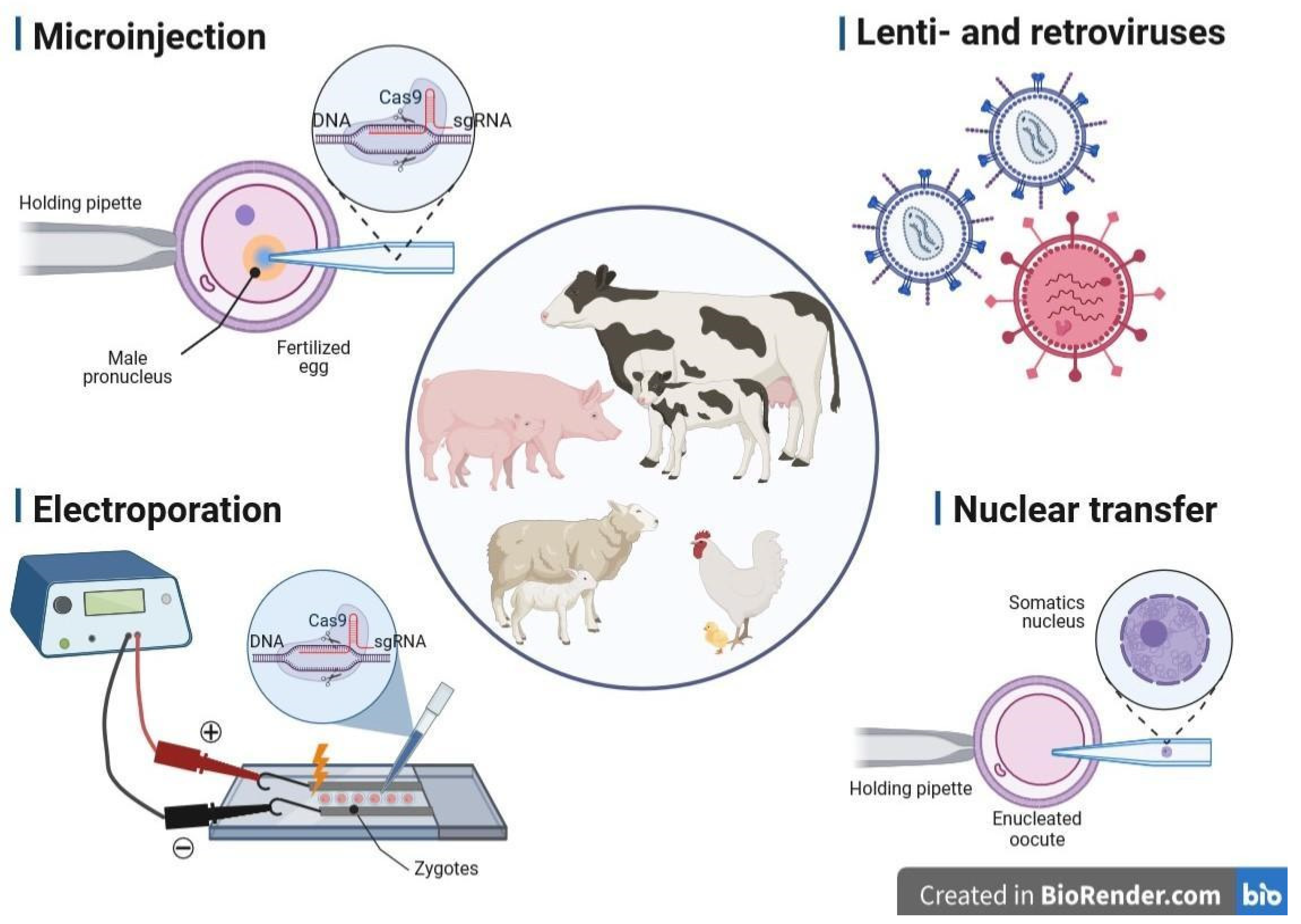
3. Methods of Gene Delivery Successfully Applied in Livestock Biotechnology
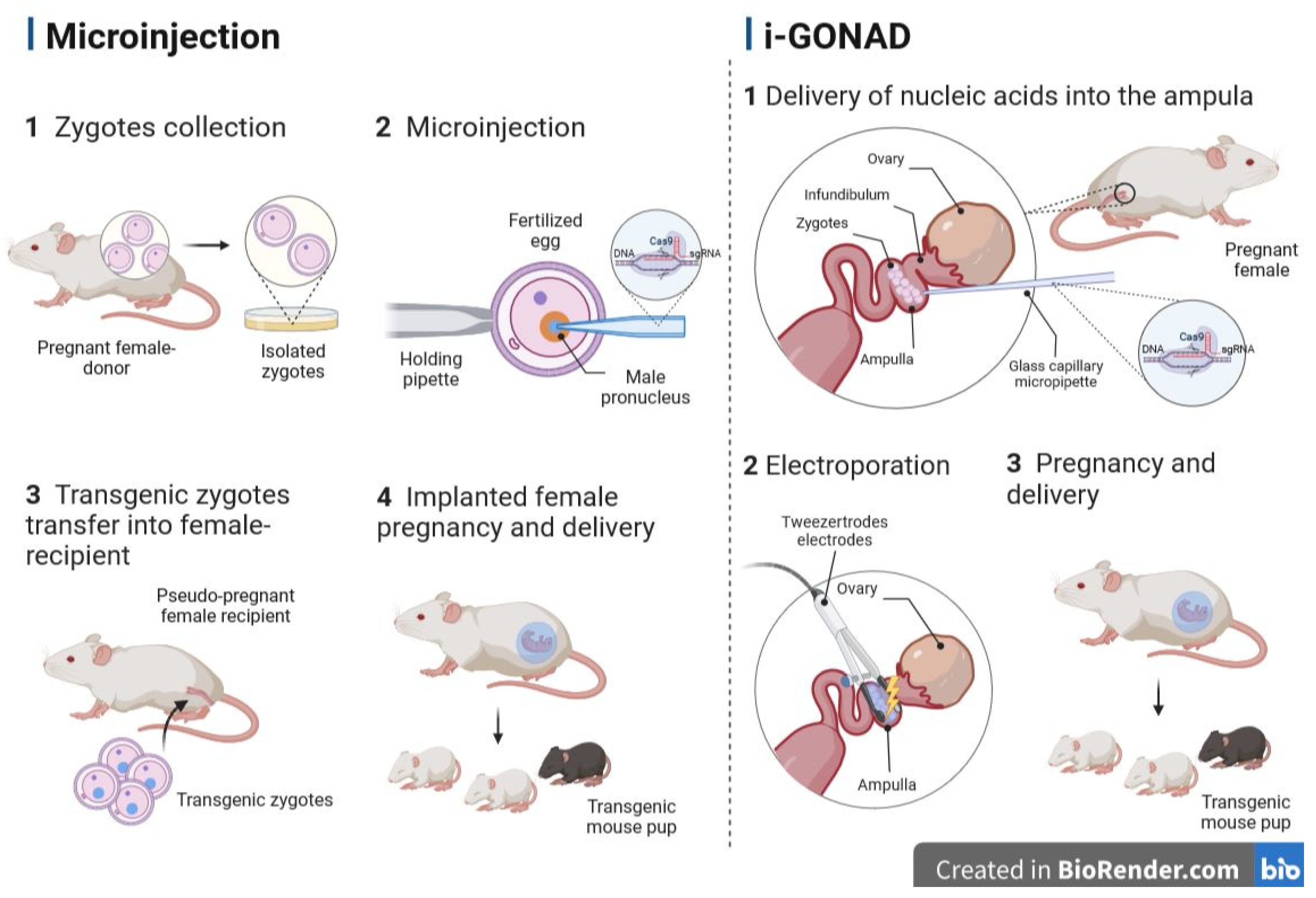
3.1. Microinjection into the Pronucleus
3.2. Retro- and Lentiviral Vectors
3.3. Electroporation of Zygotes
3.4. Transfer of Nucleus (Cloning, SCNT)
5.2. Genome Editing via Oviductal Nucleic Acid Delivery (GONAD)
5.3. Transplacental Gene Delivery (TPGD)
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Van Eenennaam, A.L.; Young, A.E. Genetic Improvement of Food Animals: Past and Future. In Encyclopedia of Food Security and Sustainability; Elsevier: Amsterdam, The Netherlands, 2018; pp. 171–180. [Google Scholar] [CrossRef]
- Singh, P.; Ali, S.A. Impact of CRISPR-Cas9-Based Genome Engineering in Farm Animals. Vet. Sci. 2021, 8, 122. [Google Scholar] [CrossRef]
- McFarlane, G.R.; Salvesen, H.A.; Sternberg, A.; Lillico, S.G. On-Farm Livestock Genome Editing Using Cutting Edge Reproductive Technologies. Front. Sustain. Food Syst. 2019, 3, 106. [Google Scholar] [CrossRef]
- Turnbull, C.; Lillemo, M.; Hvoslef-Eide, T.A.K. Global Regulation of Genetically Modified Crops Amid the Gene Edited Crop Boom—A Review. Front. Plant Sci. 2021, 12, 630396. [Google Scholar] [CrossRef] [PubMed]
- Prakash, D.; Verma, S.; Bhatia, R.; Tiwary, B.N. Risks and Precautions of Genetically Modified Organisms. ISRN Ecol. 2011, 2011, 369573. [Google Scholar] [CrossRef] [Green Version]
- Hallerman, E.M.; Bredlau, J.P.; Camargo, L.S.A.; Dagli, M.L.Z.; Karembu, M.; Ngure, G.; Romero-Aldemita, R.; Rocha-Salavarrieta, P.J.; Tizard, M.; Walton, M.; et al. Towards progressive regulatory approaches for agricultural applications of animal biotechnology. Transgenic Res. 2022, 31, 167–199. [Google Scholar] [CrossRef]
- Wells, D.J. Genetically Modified Animals and Pharmacological Research. Comp. Vet. Pharmacol. 2010, 199, 213–226. [Google Scholar] [CrossRef]
- Niemann, H.; Kues, W.A. Transgenic farm animals: An update. Reprod. Fertil. Dev. 2007, 19, 762–770. [Google Scholar] [CrossRef]
- Eriksson, S.; Jonas, E.; Rydhmer, L.; Röcklinsberg, H. Invited review: Breeding and ethical perspectives on genetically modified and genome edited cattle. J. Dairy Sci. 2018, 101, 1–17. [Google Scholar] [CrossRef]
- Van Eenennaam, A.L. Application of genome editing in farm animals: Cattle. Transgenic Res. 2019, 28, 93–100. [Google Scholar] [CrossRef]
- Niemann, H.; Kues, W.; Carnwath, J.W. Transgenic Farm Animals: Current Status and Perspectives for Agriculture and Biomedicine; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Houdebine, L.-M. Production of pharmaceutical proteins by transgenic animals. Comp. Immunol. Microbiol. Infect. Dis. 2009, 32, 107–121. [Google Scholar] [CrossRef]
- Hata, T.; Uemoto, S.; Kobayashi, E. Transplantable liver production plan. Organogenesis 2013, 9, 235–238. [Google Scholar] [CrossRef] [Green Version]
- Hryhorowicz, M.; Zeyland, J.; Słomski, R.; Lipiński, D. Genetically Modified Pigs as Organ Donors for Xenotransplantation. Mol. Biotechnol. 2017, 59, 435–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, J.; Gill, W.W. Genome Editing in Large Animals. J. Equine Vet. Sci. 2016, 41, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, Y.; Yamashita, K.; Saito, T.; Tanaka, K.; Makino, T.; Takahashi, T.; Kurokawa, Y.; Yamasaki, M.; Eguchi, H.; Doki, Y.; et al. Heparinized swine models for better surgical/endoscopic training. DEN Open 2021, 2, e64. [Google Scholar] [CrossRef]
- Tamme, R.; Laing, D.; Steinmann, W.-D.; Bauer, T. Transgenic Livestock for Food Production, Introduction. In Encyclopedia of Sustainability Science and Technology; Elsevier: Amsterdam, The Netherlands, 2012; pp. 10812–10814. [Google Scholar] [CrossRef]
- Keiser, N.W.; Engelhardt, J.F. New animal models of cystic fibrosis: What are they teaching us? Curr. Opin. Pulm. Med. 2011, 17, 478–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noble, M.S.; Rodriguez-Zas, S.; Cook, J.B.; Bleck, G.T.; Hurley, W.L.; Wheeler, M.B. Lactational performance of first-parity transgenic gilts expressing bovine alpha-lactalbumin in their milk. J. Anim. Sci. 2002, 80, 1090–1096. [Google Scholar] [CrossRef]
- Carlson, D.F.; Lancto, C.A.; Zang, B.; Kim, E.-S.; Walton, M.; Oldeschulte, D.; Seabury, C.; Sonstegard, T.S.; Fahrenkrug, S.C. Production of hornless dairy cattle from genome-edited cell lines. Nat. Biotechnol. 2016, 34, 479–481. [Google Scholar] [CrossRef]
- Young, A.E.; Mansour, T.A.; McNabb, B.R.; Owen, J.R.; Trott, J.F.; Brown, C.T.; Van Eenennaam, A.L. Genomic and phenotypic analyses of six offspring of a genome-edited hornless bull. Nat. Biotechnol. 2019, 38, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.; Zhao, X.; Zhou, J.; Li, Y.; Huang, Y.; Cai, B.; Liu, Y.; Ding, Q.; Zhou, S.; Zhao, J.; et al. Efficient generation of goats with defined point mutation (I397V) in GDF9 through CRISPR/Cas9. Reprod. Fertil. Dev. 2018, 30, 307. [Google Scholar] [CrossRef]
- Fabre, S.; Pierre, A.; Mulsant, P.; Bodin, L.; Di Pasquale, E.; Persani, L.; Monget, P.; Monniaux, D. Regulation of ovulation rate in mammals: Contribution of sheep genetic models. Reprod. Biol. Endocrinol. 2006, 4, 20. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Lin, J.; Huang, J.; Zhang, H.; Zhang, R.; Zhang, X.; Cao, C.; Hambly, C.; Qin, G.; Yao, J.; et al. Reconstitution of UCP1 using CRISPR/Cas9 in the white adipose tissue of pigs decreases fat deposition and improves thermogenic capacity. Proc. Natl. Acad. Sci. USA 2017, 114, E9474–E9482. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yu, H.; Lei, A.; Zhou, J.; Zeng, W.; Zhu, H.; Dong, Z.; Niu, Y.; Shi, B.; Cai, B.; et al. Generation of gene-modified goats targeting MSTN and FGF5 via zygote injection of CRISPR/Cas9 system. Sci. Rep. 2015, 5, srep13878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oishi, I.; Yoshii, K.; Miyahara, D.; Kagami, H.; Tagami, T. Targeted mutagenesis in chicken using CRISPR/Cas9 system. Sci. Rep. 2016, 6, 23980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, D.K.; Pinzón, C.; Huggins, S.; Pryor, J.H.; Falck, A.; Herman, F.; Oldeschulte, J.; Chavez, M.B.; Foster, B.L.; White, S.H.; et al. Genetic engineering a large animal model of human hypophosphatasia in sheep. Sci. Rep. 2018, 8, 16945. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Wang, Z.; Polejaeva, I. 40 KNOCKOUT OF GOAT NUCLEOPORIN 155 (NUP155) GENE USING CRISPR/Cas9 SYSTEMS. Reprod. Fertil. Dev. 2014, 26, 134. [Google Scholar] [CrossRef]
- Ni, W.; Qiao, J.; Hu, S.; Zhao, X.; Regouski, M.; Yang, M.; Polejaeva, I.A.; Chen, C. Efficient Gene Knockout in Goats Using CRISPR/Cas9 System. PLoS ONE 2014, 9, e106718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Program and Abstracts of the 14th Transgenic Technology Meeting (TT2017): Snowbird Resort, Salt Lake City, Utah, USA, 1–4 October 2017. Transgenic Res. 2017, 26, 1–45. [CrossRef] [PubMed]
- Fan, Z.; Yang, M.; Regouski, M.; Polejaeva, I.A. Gene Knockouts in Goats Using CRISPR/Cas9 System and Somatic Cell Nuclear Transfer. Methods Mol. Biol. 2019, 1874, 373–390. [Google Scholar] [CrossRef] [PubMed]
- Vilarino, M.; Rashid, S.T.; Suchy, F.P.; McNabb, B.R.; van der Meulen, T.; Fine, E.J.; Ahsan, S.D.; Mursaliyev, N.; Sebastiano, V.; Diab, S.S.; et al. CRISPR/Cas9 microinjection in oocytes disables pancreas development in sheep. Sci. Rep. 2017, 7, 17472. [Google Scholar] [CrossRef] [Green Version]
- Vilarino, M.; Suchy, F.P.; Rashid, S.T.; Lindsay, H.; Reyes, J.; McNabb, B.R.; van der Meulen, T.; Huising, M.O.; Nakauchi, H.; Ross, P.J. Mosaicism diminishes the value of pre-implantation embryo biopsies for detecting CRISPR/Cas9 induced mutations in sheep. Transgenic Res. 2018, 27, 525–537. [Google Scholar] [CrossRef]
- Carroll, D. Genome editing: Past, present, and future. Yale J. Biol. Med. 2017, 90, 653–659. [Google Scholar] [PubMed]
- Sun, Z.; Wang, M.; Han, S.; Ma, S.; Zou, Z.; Ding, F.; Li, X.; Li, L.; Tang, B.; Wang, H.; et al. Production of hypoallergenic milk from DNA-free beta-lactoglobulin (BLG) gene knockout cow using zinc-finger nucleases mRNA. Sci. Rep. 2018, 8, 15430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauschild, J.; Petersen, B.; Santiago, Y.; Queisser, A.-L.; Carnwath, J.W.; Lucas-Hahn, A.; Zhang, L.; Meng, X.; Gregory, P.D.; Schwinzer, R.; et al. Efficient generation of a biallelic knockout in pigs using zinc-finger nucleases. Proc. Natl. Acad. Sci. USA 2011, 108, 12013–12017. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Y.; Tian, Y.; Yu, Y.; Gao, M.; Hu, G.; Su, F.; Pan, S.; Luo, Y.; Guo, Z.; et al. Generation of mastitis resistance in cows by targeting human lysozyme gene to β-casein locus using zinc-finger nucleases. Proc. R. Soc. B Boil. Sci. 2014, 281, 20133368. [Google Scholar] [CrossRef]
- Flisikowska, T.; Thorey, I.S.; Offner, S.; Ros, F.; Lifke, V.; Zeitler, B.; Rottmann, O.; Vincent, A.; Zhang, L.; Jenkins, S.; et al. Efficient Immunoglobulin Gene Disruption and Targeted Replacement in Rabbit Using Zinc Finger Nucleases. PLoS ONE 2011, 6, e21045. [Google Scholar] [CrossRef] [Green Version]
- Geurts, A.M.; Cost, G.J.; Freyvert, Y.; Zeitler, B.; Miller, J.C.; Choi, V.M.; Jenkins, S.S.; Wood, A.; Cui, X.; Meng, X.; et al. Knockout Rats via Embryo Microinjection of Zinc-Finger Nucleases. Science 2009, 325, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lillico, S.G.; Proudfoot, C.; King, T.J.; Tan, W.; Zhang, L.; Mardjuki, R.; Paschon, D.E.; Rebar, E.J.; Urnov, F.D.; Mileham, A.J.; et al. Mammalian interspecies substitution of immune modulatory alleles by genome editing. Sci. Rep. 2016, 6, 21645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedell, V.M.; Wang, Y.; Campbell, J.M.; Poshusta, T.L.; Starker, C.G.; Krug, R.G., 2nd; Tan, W.; Penheiter, S.G.; Ma, A.C.; Leung, A.Y.H.; et al. In vivo genome editing using a high-efficiency TALEN system. Nature 2012, 491, 114–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanjana, N.E.; Cong, L.; Zhou, Y.; Cunniff, M.M.; Feng, G.; Zhang, F. A transcription activator-like effector toolbox for genome engineering. Nat. Protoc. 2012, 7, 171–192. [Google Scholar] [CrossRef] [Green Version]
- Moscou, M.J.; Bogdanove, A.J. A Simple Cipher Governs DNA Recognition by TAL Effectors. Science 2009, 326, 1501. [Google Scholar] [CrossRef]
- Method of the Year 2011. Nat. Methods 2011, 9, 1. [CrossRef] [Green Version]
- Moghaddassi, S.; Eyestone, W.; Bishop, C.E. TALEN-Mediated Modification of the Bovine Genome for Large-Scale Production of Human Serum Albumin. PLoS ONE 2014, 9, e89631. [Google Scholar] [CrossRef] [PubMed]
- Carlson, D.F.; Tan, W.; Lillico, S.G.; Stverakova, D.; Proudfoot, C.; Christian, M.; Voytas, D.F.; Long, C.R.; Whitelaw, C.B.A.; Fahrenkrug, S.C. Efficient TALEN-mediated gene knockout in livestock. Proc. Natl. Acad. Sci. USA 2012, 109, 17382–17387. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [Green Version]
- **ek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Mali, P.; Esvelt, K.M.; Church, G.M. Cas9 as a versatile tool for engineering biology. Nat. Methods 2013, 10, 957–963. [Google Scholar] [CrossRef] [Green Version]
- Hao, F.; Yan, W.; Li, X.; Wang, H.; Wang, Y.; Hu, X.; Liu, X.; Liang, H.; Liu, D. Generation of Cashmere Goats Carrying an EDAR Gene Mutant Using CRISPR-Cas9-Mediated Genome Editing. Int. J. Biol. Sci. 2018, 14, 427–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalds, P.; Zhou, S.; Cai, B.; Liu, J.; Wang, Y.; Petersen, B.; Sonstegard, T.; Wang, X.; Chen, Y. Sheep and Goat Genome Engineering: From Random Transgenesis to the CRISPR Era. Front. Genet. 2019, 10, 750. [Google Scholar] [CrossRef] [Green Version]
- Zinovieva, N.A.; Volkova, N.A.; Bagirov, V.A.; Brem, G. Transgenic farm animals: Status of the current researches and the future. Ecol. Genet. 2015, 13, 58–76. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, M.; Yamashita, Y.; Takemoto, T. Electroporation of Cas9 protein/sgRNA into early pronuclear zygotes generates non-mosaic mutants in the mouse. Dev. Biol. 2016, 418, 1–9. [Google Scholar] [CrossRef]
- O’Neil, E.V.; Brooks, K.; Burns, G.W.; Ortega, M.S.; Denicol, A.C.; Aguiar, L.H.; Pedroza, G.H.; Benne, J.; Spencer, T.E. Prostaglandin-endoperoxide synthase 2 is not required for preimplantation ovine conceptus development in sheep. Mol. Reprod. Dev. 2020, 87, 142–151. [Google Scholar] [CrossRef]
- Heo, Y.T.; Quan, X.; Xu, Y.N.; Baek, S.; Choi, H.; Kim, N.-H.; Kim, J. CRISPR/Cas9 Nuclease-Mediated Gene Knock-In in Bovine-Induced Pluripotent Cells. Stem Cells Dev. 2015, 24, 393–402. [Google Scholar] [CrossRef]
- Tan, W.; Carlson, D.F.; Lancto, C.A.; Garbe, J.R.; Webster, D.A.; Hackett, P.B.; Fahrenkrug, S.C. Efficient nonmeiotic allele introgression in livestock using custom endonucleases. Proc. Natl. Acad. Sci. USA 2013, 110, 16526–16531. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Nie, J.; Tang, Y.; He, W.; ** directly from mutant embryonic stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 4455–4460. [Google Scholar] [CrossRef]
- MacKay, G.E.; West, J.D. Fate of tetraploid cells in 4n↔2n chimeric mouse blastocysts. Mech. Dev. 2005, 122, 1266–1281. [Google Scholar] [CrossRef]
- Eakin, G.S.; Hadjantonakis, A.-K.; Papaioannou, V.E.; Behringer, R.R. Developmental potential and behavior of tetraploid cells in the mouse embryo. Dev. Biol. 2005, 288, 150–159. [Google Scholar] [CrossRef] [Green Version]
- Misra, R.P.; Bronson, S.K.; **ao, Q.; Garrison, W.; Li, J.; Zhao, R.; Duncan, S.A. Generation of single-copy transgenic mouse embryos directly from ES cells by tetraploid embryo complementation. BMC Biotechnol. 2001, 1, 12. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Talluri, T.R.; Selokar, N.L.; Hyder, I.; Kues, W.A. Perspectives of pluripotent stem cells in livestock. World J. Stem Cells 2021, 13, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Shiue, Y.; Bertolini, L.; Medrano, J.; BonDurant, R.; Anderson, G. Establishment of pluripotent cell lines from porcine preimplantation embryos. Theriogenology 1999, 52, 195–212. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, D.; Hou, Y.; Jiao, L.; Zheng, X.; Wang, W.-H. Isolation and culture of embryonic stem cells from porcine blastocysts. Mol. Reprod. Dev. 2003, 65, 429–434. [Google Scholar] [CrossRef]
- First, N.; Sims, M.; Park, S.; Kent-First, M. Systems for production of calves from cultured bovine embryonic cells. Reprod. Fertil. Dev. 1994, 6, 553–562. [Google Scholar] [CrossRef]
- Mitalipova, M.; Beyhan, Z.; First, N.L.; Hou, D.-R.; **, Y.; Nie, X.-W.; Zhang, M.-L.; Ta, N.; Zhao, L.-H.; Yang, N.; et al. Pluripotency of Bovine Embryonic Cell Line Derived from Precompacting Embryos. Cloning 2001, 3, 59–67. [Google Scholar] [CrossRef]
- Yadav, P.S.; Kues, W.A.; Herrmann, D.; Carnwath, J.W.; Niemann, H. Bovine ICM derived cells express theOct4 ortholog. Mol. Reprod. Dev. 2005, 72, 182–190. [Google Scholar] [CrossRef]
- Zhu, S.-X.; Sun, Z.; Zhang, J.-P. Ovine (Ovis aries) blastula from an in vitro production system and isolation of primary embryonic stem cells. Zygote 2007, 15, 35–41. [Google Scholar] [CrossRef]
- Behboodi, E.; Bondareva, A.; Begin, I.; Rao, K.; Neveu, N.; Pierson, J.; Wylie, C.; Piero, F.; Huang, Y.; Zeng, W.; et al. Establishment of goat embryonic stem cells from in vivo produced blastocyst-stage embryos. Mol. Reprod. Dev. 2011, 78, 202–211. [Google Scholar] [CrossRef]
- De, A.K.; Malakar, D.; Akshey, Y.S.; Jena, M.K.; Dutta, R. Isolation and Characterization of Embryonic Stem Cell-Like Cells From in vitro Produced Goat (Capra hircus) Embryos. Anim. Biotechnol. 2011, 22, 181–196. [Google Scholar] [CrossRef]
- Saito, S.; Ugai, H.; Sawai, K.; Yamamoto, Y.; Minamihashi, A.; Kurosaka, K.; Kobayashi, Y.; Murata, T.; Obata, Y.; Yokoyama, K. Isolation of embryonic stem-like cells from equine blastocysts and their differentiation in vitro1. FEBS Lett. 2002, 531, 389–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, R.; George, A.; Kamble, N.M.; Singh, K.P.; Chauhan, M.S.; Singla, S.K.; Palta, P.; Sood, T.J.; Lagah, S.V.; Sharma, A.; et al. Optimization of Culture Conditions to Support Long-Term Self-Renewal of Buffalo (Bubalus bubalis) Embryonic Stem Cell-Like Cells. Cell. Reprogramming 2011, 13, 539–549. [Google Scholar] [CrossRef]
- Koyama, S.; Kimura, T.; Ogita, K.; Nakamura, H.; Tabata, C.; Ali, K.M.A.H.N.; Temma-Asano, K.; Shimoya, K.; Tsutsui, T.; Koyama, M.; et al. Simple and highly efficient method for transient in vivo gene transfer to mid-late pregnant mouse uterus. J. Reprod. Immunol. 2006, 70, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, G.; Gurumurthy, C.B.; Wada, K.; Miura, H.; Sato, M.; Ohtsuka, M. GONAD: Genome-editing via Oviductal Nucleic Acids Delivery system: A novel microinjection independent genome engineering method in mice. Sci. Rep. 2015, 5, 11406. [Google Scholar] [CrossRef] [Green Version]
- Ohtsuka, M.; Sato, M.; Miura, H.; Takabayashi, S.; Matsuyama, M.; Koyano, T.; Arifin, N.; Nakamura, S.; Wada, K.; Gurumurthy, C.B. i-GONAD: A robust method for in situ germline genome engineering using CRISPR nucleases. Genome Biol. 2018, 19, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takabayashi, S.; Aoshima, T.; Kabashima, K.; Aoto, K.; Ohtsuka, M.; Sato, M. i-GONAD (improved genome-editing via oviductal nucleic acids delivery), a convenient in vivo tool to produce genome-edited rats. Sci. Rep. 2018, 8, 12059. [Google Scholar] [CrossRef] [Green Version]
- Hirose, M.; Honda, A.; Fulka, H.; Tamura-Nakano, M.; Matoba, S.; Tomishima, T.; Mochida, K.; Hasegawa, A.; Nagashima, K.; Inoue, K.; et al. Acrosin is essential for sperm penetration through the zona pellucida in hamsters. Proc. Natl. Acad. Sci. USA 2020, 117, 2513–2518. [Google Scholar] [CrossRef]
- Tsukamoto, M.; Ochiya, T.; Yoshida, S.; Sugimura, T.; Terada, M. Gene transfer and expression in progeny after intravenous DNA injection into pregnant mice. Nat. Genet. 1995, 9, 243–248. [Google Scholar] [CrossRef]
- Nakamura, S.; Watanabe, S.; Ando, N.; Ishihara, M.; Sato, M. Transplacental Gene Delivery (TPGD) as a Noninvasive Tool for Fetal Gene Manipulation in Mice. Int. J. Mol. Sci. 2019, 20, 5926. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Ishihara, M.; Ando, N.; Watanabe, S.; Sakurai, T.; Sato, M. Transplacental delivery of genome editing components causes mutations in embryonic cardiomyocytes of mid-gestational murine fetuses. IUBMB Life 2019, 71, 835–844. [Google Scholar] [CrossRef]
- Nakamura, S.; Ando, N.; Watanabe, S.; Akasaka, E.; Ishihara, M.; Sato, M. Hydrodynamics-Based Transplacental Delivery as a Useful Noninvasive Tool for Manipulating Fetal Genome. Cells 2020, 9, 1744. [Google Scholar] [CrossRef] [PubMed]
- Madsen, E.M.; Lindegaard, M.L.; Andersen, C.B.; Damm, P.; Nielsen, L.B. Human Placenta Secretes Apolipoprotein B-100-containing Lipoproteins. J. Biol. Chem. 2004, 279, 55271–55276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckman, D.A.; Lloyd, J.B.; Brent, R.L. Investigations into mechanisms of amino acid supply to the rat embryo using whole-embryo culture. Int. J. Dev. Biol. 1997, 41, 315–318. [Google Scholar] [PubMed]
- Rugh, R. The Mouse: Its Reproduction and Development; Oxford University Press: Oxford, UK, 1990; Volume 438. [Google Scholar]
- Kikuchi, N.; Nakamura, S.; Ohtsuka, M.; Kimura, M.; Sato, M. Possible mechanism of gene transfer into early to mid-gestational mouse fetuses by tail vein injection. Gene Ther. 2002, 9, 1529–1541. [Google Scholar] [CrossRef]
- Yamashita, M.S.; Melo, E.O. Animal Transgenesis and Cloning: Combined Development and Future Perspectives. Methods Mol. Biol. 2023, 2647, 121–149. [Google Scholar] [CrossRef]
- Galli, C.; Lazzari, G. Current applications of SCNT in advanced breeding and genome editing in livestock. Reproduction 2021, 162, F23–F32. [Google Scholar] [CrossRef]
- Morita, K.; Honda, A.; Asano, M. A Simple and Efficient Method for Generating KO Rats Using In Vitro Fertilized Oocytes. Methods Mol. Biol. 2023, 2637, 233–246. [Google Scholar] [CrossRef]
- Namula, Z.; Le, Q.A.; Wittayarat, M.; Lin, Q.; Takebayashi, K.; Hirata, M.; Do, L.T.K.; Tanihara, F.; Otoi, T. Triple gene editing in porcine embryos using electroporation alone or in combination with microinjection. Vet. World 2022, 15, 496–501. [Google Scholar] [CrossRef]
- Bevacqua, R.; Fernandez-Martín, R.; Savy, V.; Canel, N.; Gismondi, M.; Kues, W.; Carlson, D.; Fahrenkrug, S.; Niemann, H.; Taboga, O.; et al. Efficient edition of the bovine PRNP prion gene in somatic cells and IVF embryos using the CRISPR/Cas9 system. Theriogenology 2016, 86, 1886–1896.e1. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Kaneko, T. Rapid and efficient production of genome-edited animals by electroporation into oocytes injected with frozen or freeze-dried sperm. Cryobiology 2019, 90, 71–74. [Google Scholar] [CrossRef]
- Mizushima, S.; Sasanami, T.; Ono, T.; Kuroiwa, A. Current Approaches to and the Application of Intracytoplasmic Sperm Injection (ICSI) for Avian Genome Editing. Genes 2023, 14, 757. [Google Scholar] [CrossRef] [PubMed]
- Lotti, S.N.; Polkoff, K.M.; Rubessa, M.; Wheeler, M.B. Modification of the Genome of Domestic Animals. Anim. Biotechnol. 2017, 28, 198–210. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Gene | Trait | Effect | Commercial Name | References |
|---|---|---|---|---|---|
| fish | gh1 | Production trait | fast-growing salmon: 2- to 6-fold as compared to the wild-type fish | AquAdvantage | [204] |
| pig | GGTA1 | Medical use: xenotransplantation | reduces the risk of transplant rejection due to no alpha-gal sugar on cell surfaces | GalSafe | [205,206,207] |
| cattle | SLICK | Breed quality | substantially increases thermotolerance and thermoregulatory ability | PRLR-SLICK cattle | [208] |
| goat | ATryn1 | Medical use: drug production | the human ATryn1 (antithrombin-III) expressed by goats in milk | ATryn | [212] |
| rabbit | C1INH | Medical use: drug production | producing recombinant human C1 esterase inhibitor (Rhucin) in milk | Ruconest | [213,214] |
| chicken | LIPA | Medical use: drug production | express lipase A, lysosomal acid type, in eggs for long-term enzyme replacement therapy | Kanuma | [215] |
| fish | mylz2 | Fluorescent protein overexpression | overexpress GFP, YFP and RFP under a strong muscle-specific mylz2 promoter | GloFish | [217] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popova, J.; Bets, V.; Kozhevnikova, E. Perspectives in Genome-Editing Techniques for Livestock. Animals 2023, 13, 2580. https://doi.org/10.3390/ani13162580
Popova J, Bets V, Kozhevnikova E. Perspectives in Genome-Editing Techniques for Livestock. Animals. 2023; 13(16):2580. https://doi.org/10.3390/ani13162580
Chicago/Turabian StylePopova, Julia, Victoria Bets, and Elena Kozhevnikova. 2023. "Perspectives in Genome-Editing Techniques for Livestock" Animals 13, no. 16: 2580. https://doi.org/10.3390/ani13162580