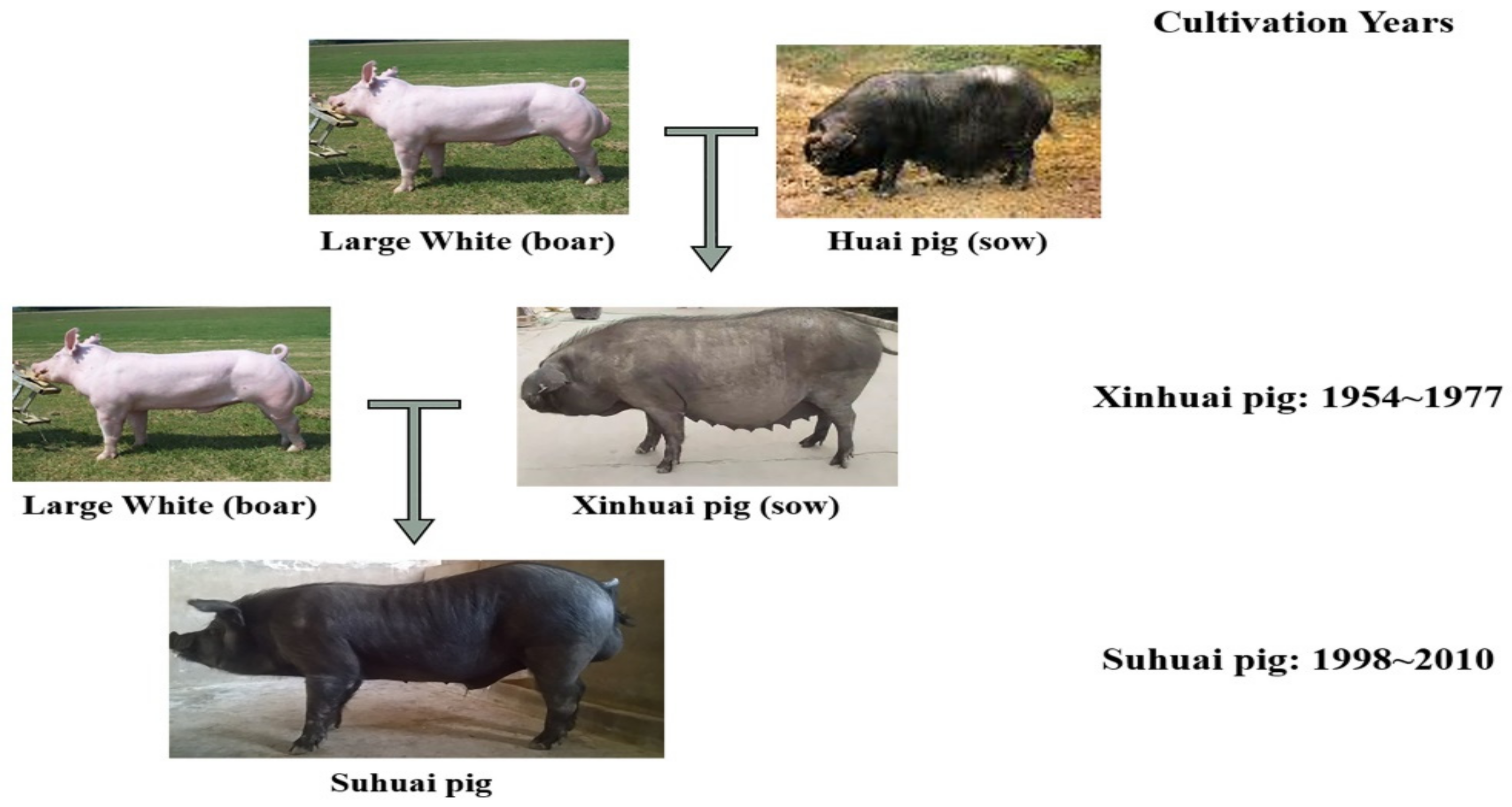
Suhuai pig is a new lean-type pig breed. Briefly, after 23 years of successive breeding of the crossbred offspring of the Large White Pig and Huai Pig, a new breed was developed, called the **
The genomic DNA was isolated from ear tissue using the standard phenol-chloroform protocol method [
30]. The DNA concentration was measured with a Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific Inc, Waltham, MA, USA) and the integrity of the DNA was checked on 1.5% agarose gel with an FR-250 electrophoresis apparatus (Furi Technology Co., Ltd., Shanghai, China). Only high-quality genomic DNAs from all samples were used for subsequent genoty**. The polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) method and the improved multiplex ligation detection reaction (iMLDR) technique [
31] were used to analyze the polymorphisms of 13 SNPs from the 12 candidate genes (
Supplementary Table S1). Among these SNPs, 11 were genotyped through iMLDR in one ligation reaction, including
LIPE (rs328830166),
IGF1 (rs341412920),
IGF1 (rs322131043),
IGF2 (g.3072G > A),
LEP (rs45431504),
LEPR (rs45435518),
MC4R (rs81219178),
RETN (rs327132149),
RYR1 (rs344435545),
SCD (rs80912566) and
UBE3C (rs81329544). Fragments covering these 11 SNPs were amplified using a multiplex of PCR reactions. The multiplex PCR reaction, performed in a 20 μL total volume, contained 1× GC-I buffer (Takara, Dalian, China), 3.0 mM Mg
2+, 0.3 mM dNTPs, 1 U of Hot-Start Taq DNA polymerase (Takara, Dalian, China), 1 μL of primer mixture, and 20 ng of genomic DNA. The PCR program was as follow: 95 °C for 2 min, 11 cycles (94 °C for 20 s, 65 °C−0.5 °C/cycle for 40 s, and 72 °C for 1 min and 30 s), 24 cycles (94 °C for 20 s, 59 °C for 30 s, and 72 °C for 1 min and 30 s), 72 °C for 2 min, and hold at 4 °C. The purification reaction, performed in a new volume, contained 10 μL of each PCR product, 5U SAP and 2U Exonuclease I at 37 °C for 1 h and at 75 °C for 15 min. The ligation reaction, performed in a 10 μL final volume, contained 1 μL of 10× ligation buffer, 0.25 μL of Taq DNA Ligase (NEB Biotechnology, Bei**g, China), 0.4 μL of 5′ ligation primer mixture, 0.4 μL of 3′ ligation primer mixture, 2 μL of purified PCR product mixture and 6 μL of double distilled water (ddH
2O). The ligation cycling program was 38 cycles (94 °C for 1 min and 56 °C for 4 min), and a hold at 4 °C. A total of 0.5 μL of ligation product was loaded into an ABI3730XL, and the raw data were analyzed using the GeneMapper 4.1 software program (Thermo Fisher Scientific Inc, Waltham, MA, USA). All of the iMLDR primers are presented in
Supplementary Table S1.
Only the polymorphisms of
FABP3 (rs1110770079) and
PHKG1 (rs697732005) were assessed using the PCR-RFLP method due to the failure of the iMLDR technique. For PCR-RFLP, the sequences for these 2 genes were obtained from the GenBank database (NLM, Bethesda, MD, USA). The primers of these 2 SNPs were designed by Primer Premier 6.0 software (Premier Biosoft International, Palo Alto, CA, USA) (
Supplementary Table S1). PCR reaction was performed in a final volume of 25 μL, containing 1 μL of the template DNA (50 ng/μL), 1 μL of each primer, 22 μL 1.3 × Taq buffer. Cycling conditions were 98 °C for 2 min; 40 cycles at 98 °C for 10 s, 57 °C for 40 s, 72 °C for 1 min; and a final extension at 72 °C for 5 min. The quality of the PCR product was analyzed by using 1.5 % agarose gel electrophoresis. Genoty** was performed by the TSINGKE Company (Nan**g, China).
2.4. Statistical Analysis
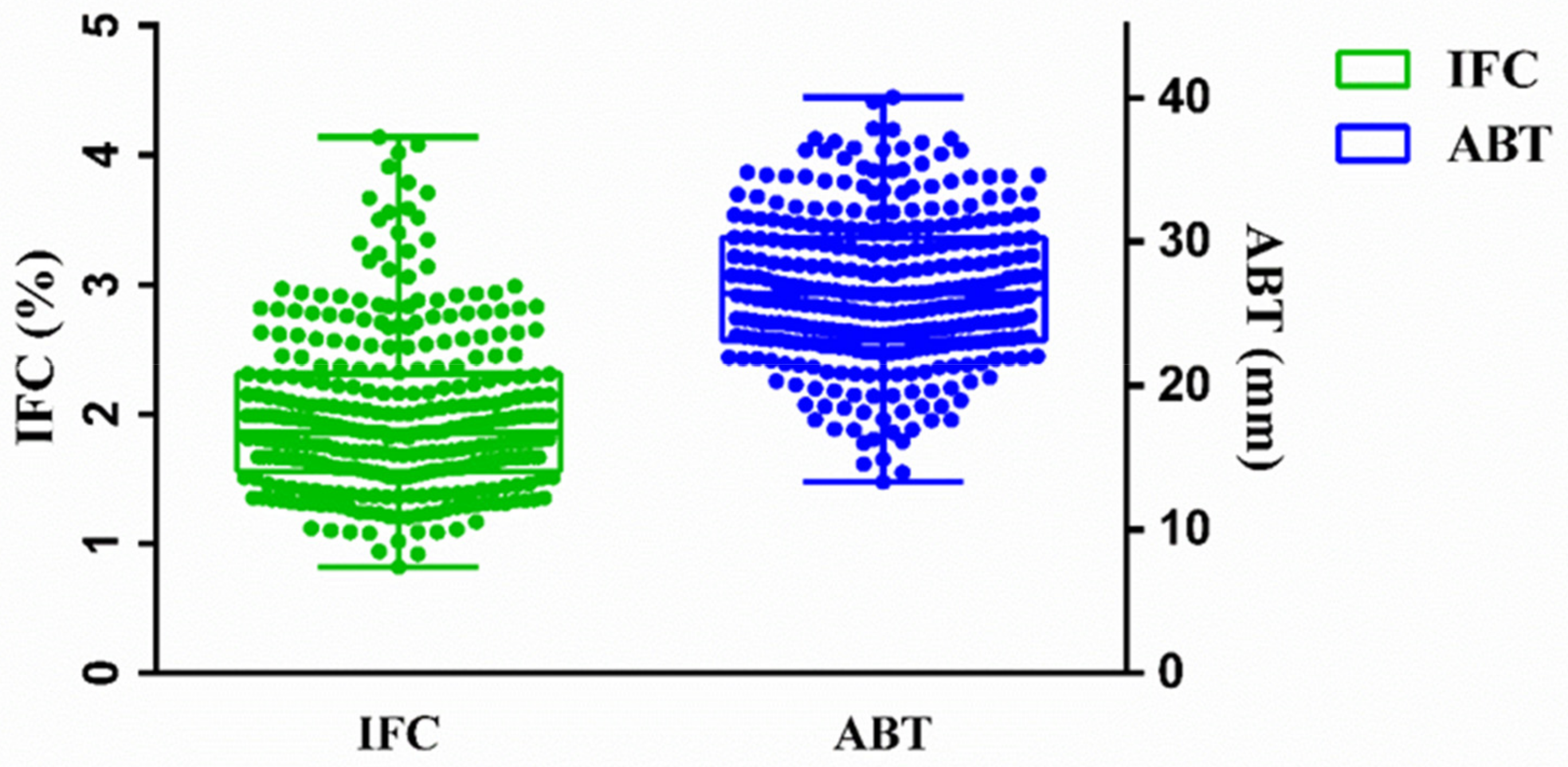
Statistical analysis of IFC and ABT of the 330 Suhuai pigs was performed using GraphPad Prism 6 (GraphPad Software Inc, La Jolla, CA, USA) with the quartile method.
Phenotypic correlations of IFC and ABT were calculated using SAS 9.2 software (SAS Institute lnc, Cary, NC, USA). To estimate genetic correlations of IFC and ABT, all the phenotypes were corrected for sex and batch as fixed effects and sample identity (ID) as a random variable using DMU software [
32].
In this study, genetic diversity indices were calculated to explore the genetic structure of all SNPs. The minimal number of genotypes used in the analysis was 3, and the total number of animals analyzed here was 327–330. The genotype and allele frequencies were calculated using Microsoft Excel 2013. The heterozygosity (He), homozygosity (Ho) and polymorphic information content (PIC) were estimated using Power Marker V3.0 software [
33].
Here, variance analyses were performed using SAS 9.2 software (SAS Institute lnc, Cary, NC, USA). Sex and batch were considered as fixed effects in the models (1) and (2), because sex and batch had significantly effects on both IFC and ABT in variance analyses. We also found that age had significant effects on IFC, and carcass weight had significantly effects on ABT. Thus, age was used as the covariate for IFC in model (1), and carcass weight was used as the covariate for ABT in model (2).
Association analysis of each SNP with IFC and ABT was performed using the PROC GLM procedure of module of SAS 9.2 software.
For IFC, the following model was applied:
For ABT, the following model was applied:
In models (1) and (2), Yijlkm represents the vector of the phenotypic value of the trait under study, μ in these 2 models is the population mean of IFC and ABT, respectively, Gi, Bj and Sk refer to fixed effects of SNPs, batch and sex, respectively, Dl represents the covariate of age for IFC, Wl is the covariate of carcass weight for ABT, Km is the random variable of kinship matrix using pedigree, and eijklm represents the random error. All statistical analyses were considered significant at p < 0.05.
4. Discussion
IFC and ABT are important economic traits for meat quality. Research on the genetic basis of these two traits has long been the focus of attention [
4,
5,
6]. Recent studies using RNA sequencing identified 5 genes that affect fat deposition [
35]. Genome-wide association studies indicated that the
CTN1 gene is associated with IFC [
36]. IFC is a relevant trait for high-quality meat products [
37], but high BF thickness leads to a decrease in lean meat percentage [
38]. Therefore, this study aimed to find molecular markers that are significantly associated with IFC, but do not change ABT of Suhuai pigs. A total of 330 Suhuai pigs were measured for IFC and ABT. The CVs for IFC and ABT were high, perhaps due to the fact that the Suhuai is a new synthetic hybrid breed of pig, and there was a difference in the age and carcass weight of this population. The mean of IFC was low, but compared with Large White, Suhuai pigs had higher IFC and BF. Phenotypic and genetic correlations between IFC and ABT were moderate. This suggests that it should be possible to increase IFC without changing BF thickness. Suzuki et al. [
12] reported that the phenotypic correlation between IFC and ABT was moderate, similar to our results.
In the present study, 13 SNPs from 12 previously reported candidate genes for IFC were genotyped, and these SNPs were polymorphic in Suhuai pigs. Among these SNPs, low polymorphisms of the alleles
IGF1 (rs322131043),
LEP (rs45431504),
LEPR (rs45435518),
RETN (rs327132149) and
RYR1 (rs344435545) were observed. These results indicated that there was a strong selection pressure on several traits that were associated with SNPs in this study, such as growth rate, yield of lean meat, BF thickness and so on. The lower values of PIC and He in the
IGF1 (rs322131043),
LEP (rs45431504),
LEPR (rs45435518) and
RETR (rs327132149) provided additional evidence for the strong selection pressure. For example, the
RYR1 (rs344435545) SNP is a missense mutation that causes a change in protein (Arg
615→Cys
615), and this mutation results in muscle dysfunction and finally porcine stress syndrome (PSS) [
39]. This disease may cause pigs to produce pale soft exudative meat (PSE) [
22]. Therefore, breeders have been working hard to eliminate deleterious alleles in recent decades.
Association analysis of the 13 evaluated SNPs with IFC and ABT was performed. Associations of all SNPs with IFC were not significant, except for the
FABP3 (rs1110770079) SNP. These 13 SNPs did not associate with ABT of Suhuai pigs; the possible reasons were that some of these genes were not causal genes for BF thickness, such as
PHKG1,
RETN,
SCD, and
UBE3C. The
FABP3 gene is one member of the fatty acid binding protein family, which plays a critical role in intracellular fatty acid transport by binding lipids and regulating metabolic homeostasis [
40]. The
FABP3 gene might be responsible for IFC and is often regarded as a candidate gene [
13]. Chen et al. [
41] found that the
FABP3 (HinfI) SNP was associated with IFC in both Yanan (
p < 0.001) and DLY (
p < 0.05) pigs, but did not significantly affect the BF thickness, strikingly similar to the results in our study. Although the rs344435545 SNP in the
RYR1 gene is likely to cause an increase in the incidence of porcine stress syndrome, there is a strong correlation between IFC and
RYR1 gene expression level [
42] and as this mutation can cause a decrease of BF thickness, it has been considered as an example of balanced selection [
43]. The
SCD (rs80912566) SNP could affect the fatty acid composition and IFC within the Duroc population [
18]. Moreover, the
SCD gene was identified as a candidate gene related to IFC between pigs with high and low IFC [
44]. The result from Henriquez-Rodriguez et al. [
45] showed that the
SCD (rs80912566) SNP was associated with fat composition but not with fat content. Their results were consistent with our study. It is worth noting that the results in
Table 3 revealed a tendency (
p < 0.1) of association between the
RYR1 (rs344435545) and
SCD (rs80912566) with IFC. We speculate that these two SNPs have small effects on IFC or these may be only in linkage disequilibrium with the causative mutation of IFC.
The
IGF1 gene affects the regulation of adipogenesis. Several results suggested a degree of positive correlation between the
IGF1 gene expression and adipocyte content [
15], and within the QTLs that affect IFC and BF thickness. Similar to the
IGF1 gene, the
LEP and
LEPR genes also play a role in adipogenesis, and several studies have found that
LEP (rs45431504) and
LEPR (rs45435518) are polymorphic and could significantly influence IFC and BF thickness [
46,
47,
48]. The
RETN gene is located on SSC2, and
RETN (rs327132149) was significantly associated with the abdominal fat weight, BF thickness and IFC [
21,
49]. On the contrary,
IGF1 (rs322131043),
LEP (rs45431504),
LEPR (rs45435518) and
RETN (rs327132149) were not significantly associated with IFC and ABT in our study and might be strongly selected for these 3 genes in Suhuai population, resulting in low polymorphisms in these SNPs.
The
LIPE gene has long been considered as a candidate gene that could affect IFC deposition due to resolving fat [
14,
50]. Burgos et al. [
16] reported that
IGF2 (g.3072G > A) could affect pig carcass traits and IFC in Large White×Landrace populations. On the contrary, Aslan et al. [
51] found that the
IGF2 (g.3072G > A) SNP may not affect IFC in Pietrain, Duroc and Large White populations. Supakankul et al. [
23] reported that these 2 porcine
UBE3C polymorphisms (rs81329544 and rs32466023) were associated with IFC and fatty acid composition. The
UBE3C gene is considered a potential candidate gene for fat deposition in muscle because of its location on SSC18, near the QTLs for IFC and FA composition [
52]. The
PHKG1 gene is related to glycolysis potential and could affect pork quality. A previous study indicated that there was a point mutation (rs330928088) in a splice acceptor site of intron 9 in the
PHKG1 gene. This point mutation gave rise to the 32 bp deletion in the open reading frame (ORF) and generated a premature stop codon [
20]. However, the
PHKG1 (rs330928088) SNP is not polymorphic and did not cause the 32 bp deletion of Suhuai pigs. The SNP (rs697732005) in front of this
PHKG1 (rs330928088) SNP is polymorphic, although it was not associated with IFC in this study. The
MC4R gene has only one exon, located in the QTL for ABT on SSC1, and plays an important role in the regulation of energy homeostasis [
53]. Several reports found that the
MC4R gene could be associated with fat mass in humans [
54] and IFC in Hu sheep [
55]. Lyadskiy et al. [
56] found no significant difference between the spinal fat thickness and the
MC4R (rs81219178) SNP, similar to our results. Nevertheless, there was no significant difference in IFC between the three genotypes of the
MC4R (rs81219178) in pigs [
19]; this result is consistent with our study. The fact that the above-mentioned five SNPs were not significantly associated with IFC and ABT might be due to genetic heterogeneity. After all, Suhuai pig is a hybrid population containing Huai pig (25%) and Large White (75%).
These 13 SNPs of the 12 genes did not associate with ABT in Suhuai pigs. The possible reasons were that some of these genes were not causal genes for BF thickness, and other candidate genes of BF thickness may have heterogeneity between breeds. Finally, the FABP3 (rs1110770079) SNP was the genetic marker we were looking for, which could improve IFC without increasing BF thickness.




{kind=link}
{kind=link}